



第17章 压电效应在生物基本结构中的生理作用
1. 生物基本单元中的压电效应
1.1. 氨基酸
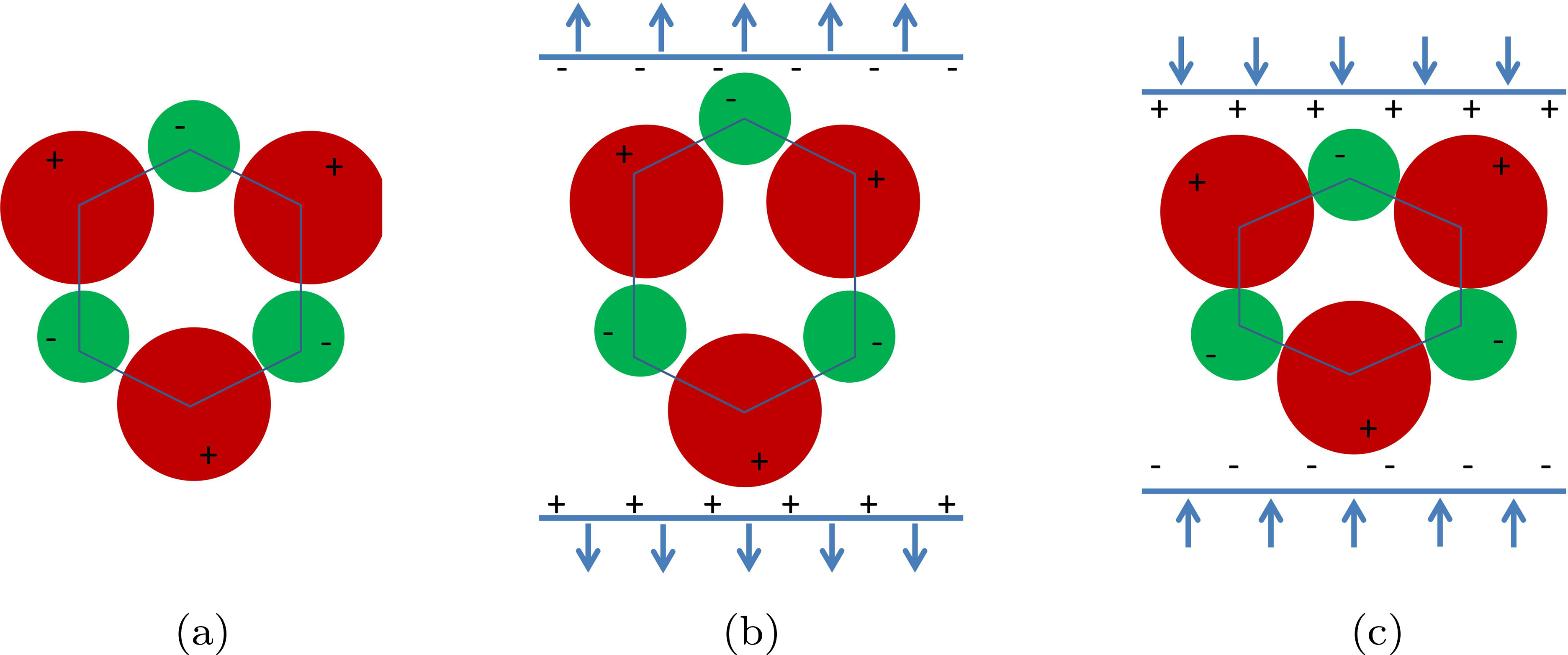
氨基酸是生物学的基本元素。它们由通过肽键连接的简单化合物组成,这些化合物串联在一起形成肽或多肽。如果氨基酸的组装形成一种生物功能肽,则称之为蛋白质。氨基酸晶体可从饱和水溶液中生长,并通过X射线衍射和核磁共振技术确定其晶体结构。由于氨基酸是手性实体,它们属于对映体对称群。在赫尔曼‐莫甘符号中,这些群可以列为1–4、6、222、32、422、622、23和432群。根据定义,属于这些对称群的氨基酸晶体没有反演中心。除 α‐甘氨酸为特例外(α‐甘氨酸属于非手性对称群2/m),20种蛋白质氨基酸中的19种应表现出非中心对称晶体的性质,即压电效应。
1970年,瓦西列斯库首次报道了氨基酸粉末中的压电共振现象。后续的研究主要集中在氨基酸及其化合物的晶体上。在这些实验中,核四极共振波谱仪向样品施加高频电压脉冲,随后检测并测量样品中由电压感应产生的振动。该仪器的灵敏度据报相当于石英压电响应的 10⁻⁴。在大多数报告中,氨基酸的压电响应以任意单位表示。然而,在一项研究中,L‐精氨酸磷酸盐的压电 d₂₅系数被测定为 8.6 皮库仑/牛顿。
氨基酸的压电特性似乎与固态材料一样,受相同的晶体结构要求支配。如图1所示,属于非中心对称对称群的氨基酸应表现出压电效应。
然而,这一理论因在 α‐甘氨酸和外消旋氨基酸中观察到压电效应而受到质疑。这些问题已被解决。在 α‐甘氨酸中观察到的压电行为可归因于其环境形态 γ‐甘氨酸的污染,后者是一种已知的压电材料。理想情况下,左旋(L)或右旋(D)氨基酸应具有手性,尽管对于属于非中心对称群mm2的DL‐丙氨酸等情形,DL型氨基酸仍可能表现出压电效应。另一方面,氨基酸中缺乏压电效应的情况对其对称性应支撑压电效应,这对理论提出了挑战。实验装置无法考虑诱导弹性振动的阻尼效应,这可能是造成差异的原因。
1.2. 脱氧核糖核酸
脱氧核糖核酸是所有生物体的遗传蓝图,负责编码发育和功能的指令。脱氧核糖核酸样品中压电效应的存在再次表明,该现象在生物学各个层面都十分普遍。Duchesne et al. 首次报道了脱氧核糖核酸中的压电效应。实验涉及将样品置于超再生振荡器的电容器中。在30兆赫处观察到的吸收线归因于由逆压电效应在DNA分子内部引发的机械振动。
尽管这些早期实验主要是定性的,但它们令人振奋,暗示了生物压电性是一种普遍存在的生物现象。福田等人利用基于正压电效应的技术,对小牛胸腺脱氧核糖核酸的取向膜进行了定量测量。样品通过轧制脱氧核糖核酸纤维直至其融合成膜而制备。研究了剪切应力作用下取向DNA薄膜的极化情况,在几种不同的温度下。据报道,小牛胸腺脱氧核糖核酸的一个典型值为 10⁻¹⁰ cgs esu(∼0.003 pC/N),低于迄今为止报道的蛋白质的数值。相对较低的压电响应归因于结晶度较低,这一点通过X射线衍射测量得到了证实。结晶度低是由于脱水过程造成的,而脱水过程是减少样品湿气引起的导电性所必需的。脱氧核糖核酸中压电效应的根本机制归因于DNA分子碱基中极性基团的应力诱导取向。
许多聚合物(包括聚偏氟乙烯(PVDF))的压电特性受样品拉伸、轧制或拉伸程度的影响。脱氧核糖核酸薄膜也是如此。在一项研究中,从小牛胸腺提取的脱氧核糖核酸样品被制备成薄膜,并被拉伸至原始长度的1.5到2.5倍。拉伸比为1.5的样品表现出最大压电系数为 −3.3 × 10⁻⁸ cgs esu(∼ −1.09 pC/N),而将拉伸比增加到2.5时,压电系数上升至 −10.2 × 10⁻⁸ cgs esu(∼ −3.37 pm/N)。以这种方式拉伸薄膜可使分子排列更加整齐,从而增强压电响应。
已知相对湿度会影响生物材料的压电特性。为了理解湿度对脱氧核糖核酸样品的影响,研究了由聚合三文鱼制备的取向DNA薄膜的正压电效应和逆压电效应。DNA钠盐可存在于三种不同的构型中,具体取决于盐含量和相对湿度。为探究湿度对脱氧核糖核酸样品压电效应的影响,实验中保持盐含量恒定(按重量计6–7%),并改变湿度。当相对湿度高于85%时,脱氧核糖核酸样品呈现B螺旋构型;在75%至85%之间时,样品具有A螺旋构型;当相对湿度低于55%时,样品呈无序状态。令人惊讶的是,研究发现含水量较低的脱氧核糖核酸样品仍表现出压电的特性。此前人们认为这些样品的无序结构会使其不具备压电性。单链DNA样品表现出与脱水样品类似的结果。通过这一对比可以提出变性DNA中的压电效应源于主链的应力诱导取向。对于水合双链DNA样品,其结构因水分子吸收而稳定。有序DNA样品中压电效应的起源归因于含有CO和NH极性基团的嘌呤和嘧啶碱基的偶极子。在水合DNA样品中,这些极性基团的重新取向以及主链的重新取向表现为压电效应。
1.3. 磷脂
脂质在细胞膜结构中起着重要作用,作为细胞与其环境之间的柔性屏障,并为膜蛋白提供发挥作用的平台。众所周知,包括磷脂、氨基酸和蛋白质在内的生物分子均表现出手性。然而,生物物质的手性常常被忽视。相反,液晶的手性在技术应用方面的重要性已得到认可。通过将磷脂与液晶同样看待进行研究,人们发现了磷脂中的压电效应。
液晶态同时表现出取向有序和流动性。液晶的这些非凡特性使其具有大量应用,最显著的是在液晶显示器中。在液晶领域内存在两大主要类别,向列相和近晶相。向列相液晶的分子不具有任何位置有序;然而它们倾向于平行于一个共同的指向矢轴排列。近晶相液晶的结构更为有序,由具有明确层间距的取向分子层构成。如果分子长轴指向垂直于层轴的方向,则称为 SmA相,而倾斜排列则称为 SmC相。液晶以这种方式排列的倾向使其具有各向异性;其电学和光学性质因相对于指向矢的测量方向不同而有所差异。
手性向列相和近晶相晶体也存在。它们的手性是在液晶中观察挠曲电性、铁电性和压电效应所必需的不对称条件的关键因素。迈耶是第一个发现液晶与其低对称性相关潜在特性的联系的人,他发表了关于液晶中存在压电效应的论点,并在1969年通过实验证实。
对磷脂中可能存在的压电效应感兴趣的研究人员注意到手性液晶与手性磷脂之间存在显著的相似性;两者都有可能表现出压电效应,但均不具有传统固态压电材料的典型外观。关于磷脂的大量理论和实验工作都是基于已有的液晶物理学知识。事实上,液晶学家雅克利最先提出假设:D型和L型构型的磷脂样品会表现出压电效应,而外消旋磷脂则不会。一项针对合成磷脂(例如二棕榈酰磷脂酰胆碱,DPPC)的研究证实,手性是磷脂基压电效应的核心;在其D型(D‐DPPC)和L型(L‐DPPC)构型中观察到显著的压电效应,而其外消旋对照(DL‐DPPC)则几乎无响应。
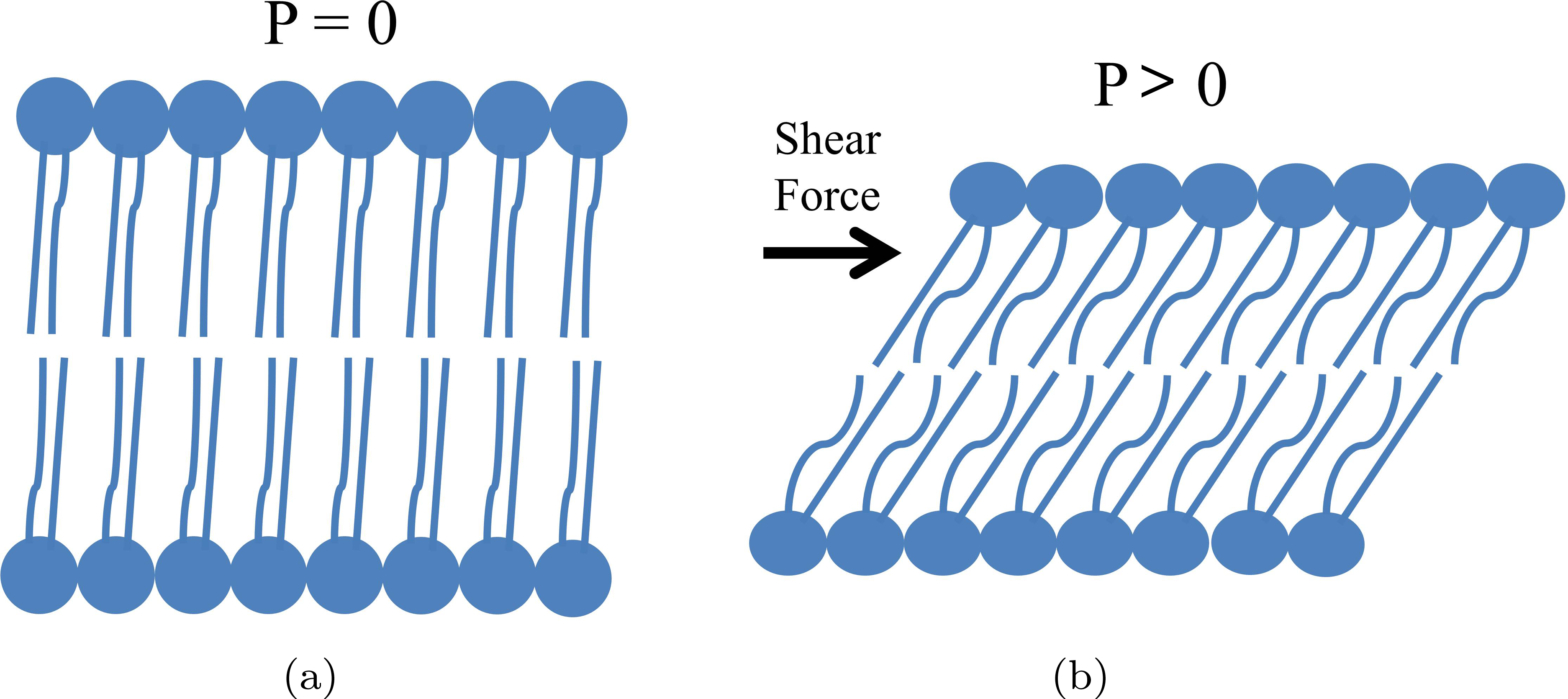
从液晶的角度来看,磷脂双分子层可被视为具有D∞对称性的 SmA*相,或具有C2对称性的SmC*相;这两种相位在剪切应力作用下均应表现出压电效应,如图2所示。对SmA*相磷脂施加剪切力会使分子发生5度倾斜,从而使净极化方向垂直于倾斜平面。对 SmC*相磷脂双分子层施加剪切应力会扭曲极化螺旋的旋转对称性,并引发压电响应。实验已证实了这一点。将磷脂样品置于两个基底之间并施加剪切应力,这导致分子产生5度倾斜,并产生300 nC/cm²的极化。
1.4. 纤维蛋白
以各种形式存在的纤维状蛋白质在细胞和组织中发挥着重要作用,支撑着多种不同的结构需求,包括机械稳定性、支撑、刚度和弹性。除了机械特性外,纤维蛋白还表现出机电性能。压电效应已在胶原蛋白、角蛋白、丝素蛋白和弹性蛋白中被观察到。由于胶原蛋白中的压电性可能在骨压电性中具有生理作用,因此相较于其他纤维蛋白,胶原蛋白的压电性得到了更广泛的研究。因此,我们将以胶原蛋白中的压电性机制为例进行综述,作为其他纤维蛋白的代表性案例。
胶原蛋白具有复杂的层级结构,如图3所示。在最低的层级上是胶原分子,其由三条多肽链缠绕在一起形成右手三重螺旋。这些链内的区域包含重复的甘氨酸–X–Y氨基酸基序,其中任何氨基酸都可以作为X和Y。胶原分子聚集在一起形成原纤维,每个分子与其相邻分子平行排列,并以交错方式排列,使得每个分子在原纤维中位于其相邻分子上方或下方约64–67纳米处。这种偏移赋予原纤维其特有的重复D带结构。胶原原纤维的横截面视图显示,胶原分子在原纤维内呈准六边形排列。
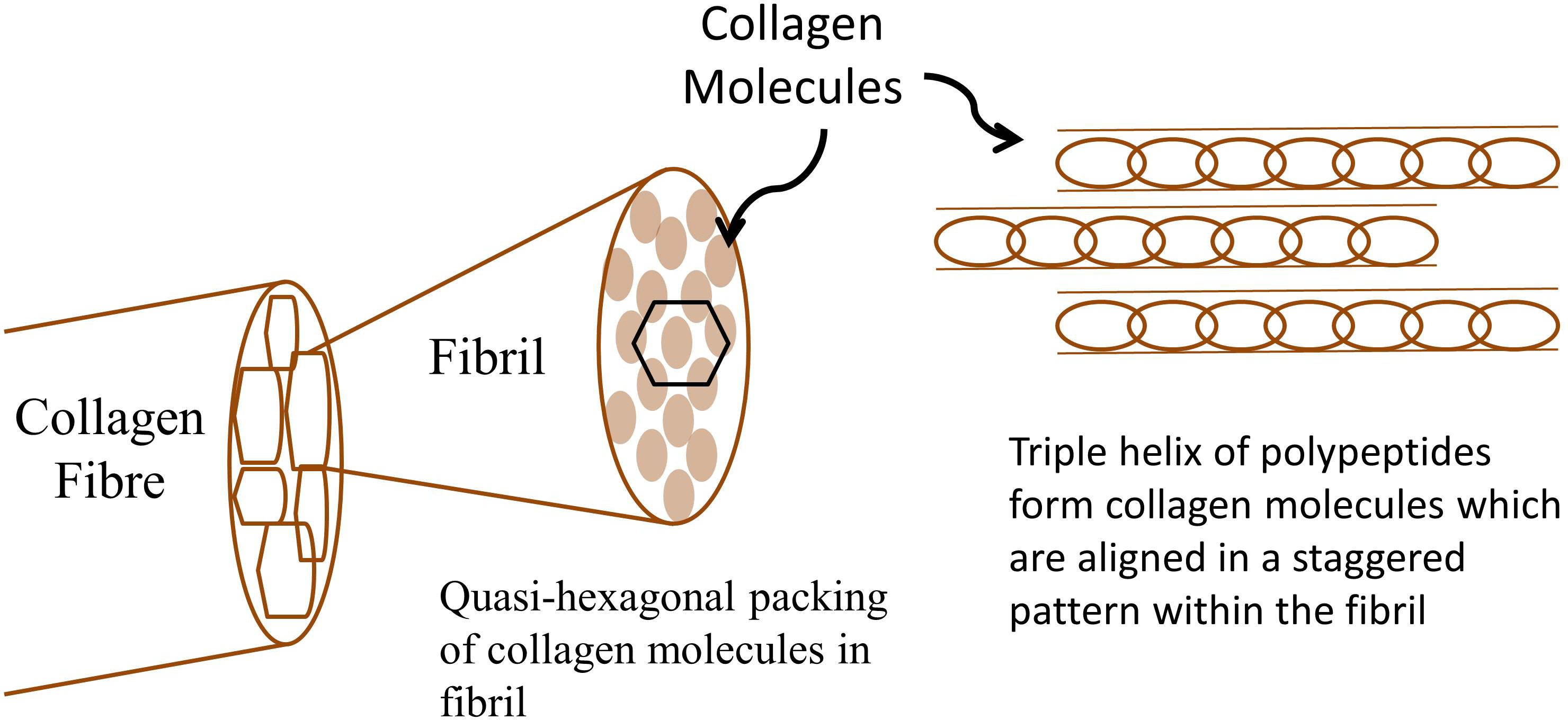
其结构的复杂性使得难以精确确定胶原蛋白中压电效应的起源。从早期对跟腱中胶原蛋白压电效应的研究来看,原纤维的高度取向性和结晶性质被认为与压电效应相关。胶原蛋白分子呈准六边形排列,可用对称群C6描述。在此对称群中,仅预期存在剪切压电性。看来,压电效应的起源可能是胶原多肽链中的氢键在受到剪切应力或电场作用时产生的诱导极化或位移。胶原蛋白中的剪切压电效应(d₁₄)测得值为 d₁₄ 8 × 10⁻⁸ cgs.esu(∼ 2.64 pC/N)。已成功构建基于胶原蛋白的留声机拾音器,展示了将胶原蛋白用作机电换能器的潜力。
压电响应力显微镜(PFM)是一种由原子力显微镜(AFM)改进而来的技术,用于研究逆压电效应。由于该技术能够在纳米尺度上探测机电性能,因此特别适用于在不同层级上研究压电效应。该技术已成功用于证明单个胶原蛋白纤维表现出剪切压电性。在纳米尺度上测得的剪切压电系数(∼ 1 pm/V)与在块体样品中测得的系数处于同一数量级。
2. 溶菌酶中的压电效应
对生物基本单元中压电效应潜在机制的全面理解仍然不足。为解决这一问题,我们特别关注蛋白质压电性研究,将其视为生物压电性研究中缺失的关键环节。在层级结构中,蛋白质连接了简单的生物组分(氨基酸)与大型复杂系统(如骨)。迄今为止,关于蛋白质压电性的研究完全集中在纤维状类型上,而忽视了球状和跨膜类型。
我们的研究聚焦于溶菌酶,这是一种存在于唾液、泪液以及大量存在于蛋清中的球状蛋白质。其主要功能是通过攻击细菌的保护性肽聚糖细胞壁来抵御细菌感染。在许多方面,溶菌酶是进行压电性案例研究的理想蛋白质:它分子小且稳定,已被广泛应用于衍射技术研究,并且能够结晶成不同的对称群。最后一点至关重要,使我们能够确定蛋白质压电性是否符合基于晶体学要求的压电性的经典描述。
压电响应力显微镜已成为在纳米尺度上对样品进行压电测量的有力工具。该技术基于逆压电效应,通过原子力显微镜悬臂施加偏压,以在压电样品中诱导形变。这些形变由同一探针在扫描样品表面时检测,并通过灵敏的激光‐光电二极管装置进行监测。
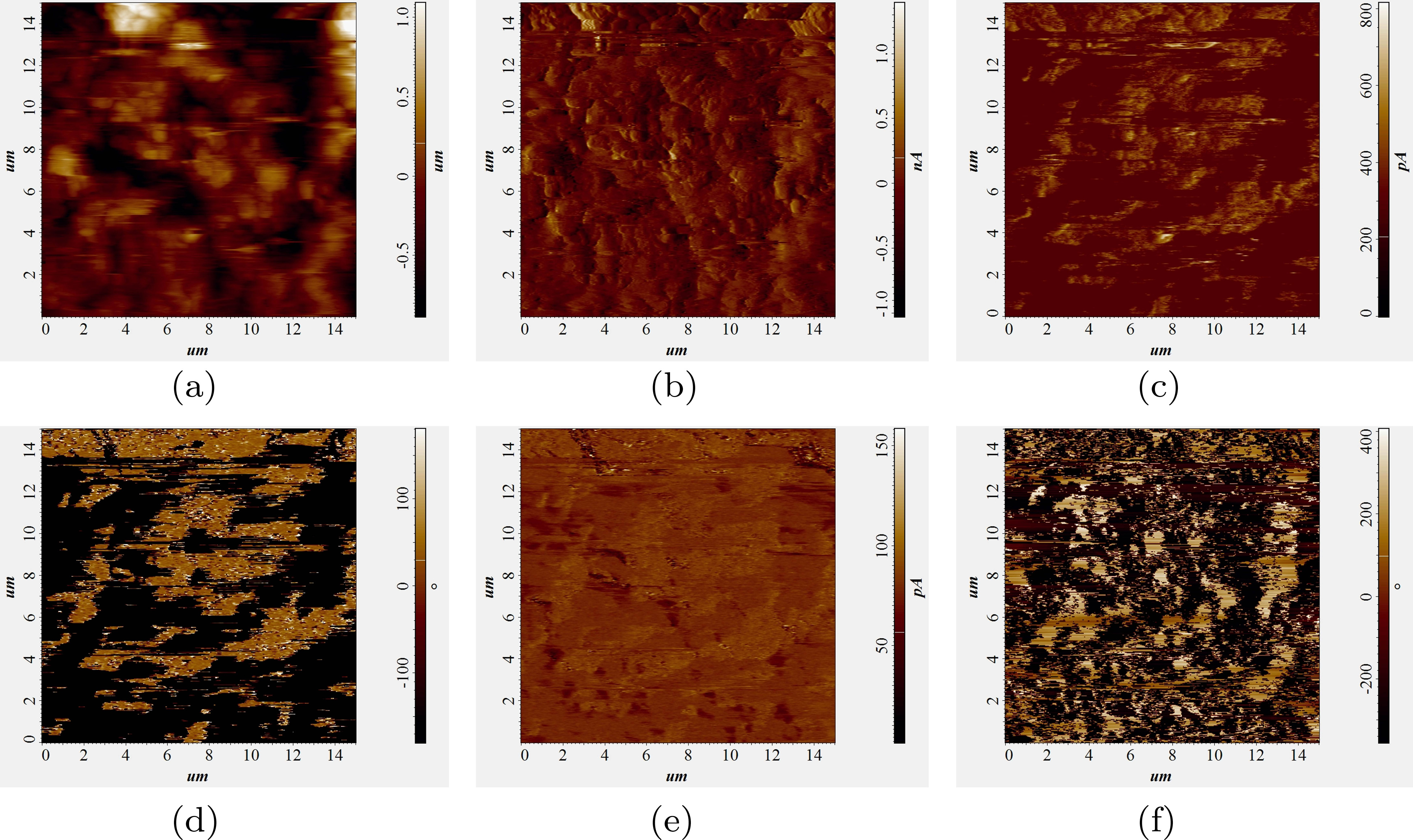
在我们对溶菌酶的研究中,PFM(压电力显微镜)非常有用,因为它使我们能够直接对单个蛋白质晶体施加电压,并确定其是否具有压电性,以及压电性的程度如何。此外,PFM能够同时监测面外和面内形变。这一特性在研究属于点群2的单斜溶菌酶晶体时尤为有利。从经典压电观点来看,此类晶体可能同时表现出横向和纵向的压电分量。图4展示了三次连续PFM扫描的结果;第一次扫描对 5 × 5 µm区域施加了10 V电压,第二次施加了5 V对一个 10 × 10 µm区域施加偏压,第三个在前两个周围的 15 × 15 µm扫描区域内施加5 V。第三次扫描显示了前两次扫描中施加偏压的影响,表明该蛋白质不仅具有压电性,而且是可极化的。存在面外和面内压电响应的证据。单斜溶菌酶晶体的压电系数估计为1 pm/V。
第17章 压电效应在生物基本结构中的生理作用(续)
3. 生理学上重要的压电效应的特殊案例
3.1. 骨
骨是一种特化的系统,不仅作为保护重要器官的骨骼框架,还为骨髓生成血细胞提供环境。它是一个动态的结构,不断经历塑造和重塑过程。骨会适应施加于其上的机械应力,这一现象由沃尔夫定律描述。据传闻,骨负荷与骨密度之间存在相互关系,广为接受。宇航员在长时间处于低重力环境后,由于骨骼承受的负荷较低,会出现骨密度下降的情况。相比之下,运动员的骨密度高于平均水平,因为他们的骨骼在训练过程中经常受到规律的负荷刺激。从临床角度来看,沃尔夫定律似乎有一定道理,但该定律本身过于简单化。
骨重塑通过成骨细胞和破骨细胞的共同作用得以实现,其中破骨细胞吸收受损的骨组织,而成骨细胞生成新的骨组织。骨如何感知负荷变化从而激活这些细胞?20世纪50年代,人们热衷于提出骨和胶原蛋白中存在压电效应,认为这是骨适应机械应力能力的支持机制。该机制十分简单:当机械应力作用于胶原纤维时,通过压电效应产生电势梯度,进而触发骨再生细胞的活动。
然而,越来越多的证据对骨压电性在骨的应变诱导电位中所起作用的程度提出了质疑。首先,关于骨压电性的大部分认识都来自于干燥骨的实验。而活体骨是湿润的。对湿骨进行的实验表明,压电效应显著减弱甚至不存在。其次,诱导电位的弛豫时间过长,不可能源自压电效应。第三,应变诱导电位具有与黏度相关的特点,这一事实与流动电位概念相符,但不符合压电效应。这些及其他论据 increasingly 指向流动电位是一种更具说服力的机制。
压电效应是否参与骨的生理过程?Marion et al. 进行了一项有趣的研究,以确定压电效应是否能在骨中引发生理功能。将具有压电性和非压电性的聚偏氟乙烯(PVDF)植入大鼠体内,并监测骨形成程度。压电性 PVDF 的植入 consistently 导致比非压电性植入更显著的骨形成。此类实验使得人们难以完全排除压电效应潜在贡献的可能性。
压电效应。Ahn et al. 提出了一种双重机制,涉及流动电位和压电效应。他们假设,压电效应可能通过机械诱导表面电位变化来影响泽塔电位(从而影响流动电位)。
3.2. Prestin蛋白
哺乳动物听觉系统是进化过程中一个引人入胜的成功范例。在人类中,听觉系统可在很宽的频率范围(20赫兹至20千赫兹)内运作,能够检测亚纳米级声振动。最近,一种名为prestin的跨膜蛋白被确定为听觉过程的关键组分。如果Prestin蛋白分子通过某种方式被降解或消除,就会观察到严重的听力下降。引人注目的是,压电效应被认为是Prestin蛋白在哺乳动物听觉中发挥作用的机制。为了理解Prestin蛋白在听觉中的作用,我们首先概述声音在听觉过程中的感知方式。
声波通过外耳传递,经由一系列中耳听小骨传导至内耳的听觉部分——耳蜗,耳蜗内含有听觉感受器柯蒂氏器。柯蒂氏器包含两种不同类型的细胞:内毛细胞(IHCs)和外毛细胞(OHCs)。内毛细胞(IHCs)作为感觉受体,将听觉信息传递给大脑。外毛细胞则对声音进行放大。当声波进入耳蜗时,附着于外毛细胞(OHCs)顶端的纤毛束发生偏转。这种偏转会激活外毛细胞(OHCs)中机械敏感离子通道的开闭,导致细胞膜两侧的电位反复变化。外毛细胞(OHCs)的一个显著特征是其能响应细胞膜电位的变化而发生构象长度变化:当细胞去极化时缩短,而在超极化时则伸长。这种电驱动运动性被称为电致运动性。因此,外毛细胞(OHCs)表现出类似于逆压电效应的行为。外毛细胞(OHCs)的压电系数极为显著,据报道为 20 µC/N,比最优的人工压电材料高出四个数量级。
Prestin蛋白已被确定为负责外毛细胞电致运动性的马达蛋白。在一项关键研究中,Liberman等生成了prestin基因敲除小鼠。从这些基因敲除小鼠中分离的外毛细胞的电致运动性显著降低。相应地,声音敏感性降低了100多倍。这些发现表明,prestin分子的降解会导致严重的听力下降。然而,基因敲除小鼠外毛细胞的机械特性发生了显著改变。去除prestin引起的细胞刚度变化可能对所观察到的听力损伤有所贡献。为了进一步研究,Dallos et al. 创建了一种基因敲入小鼠,其外毛细胞含有突变型prestin。该突变型prestin不能支持电致运动性,但维持了细胞正常的机械特性。在基因敲入小鼠中观察到了严重的电致运动性降低和听力损伤。这些研究对于确立基于prestin的外毛细胞电致运动性在耳蜗中的声音放大和频率调谐中的必要性具有关键作用。
OHC的电致运动性与压电效应非常相似。但这种现象是否属于经典意义上的压电效应呢?电致运动性满足压电效应的一些关键条件:它涉及机电耦合,且该效应具有可逆性。然而,目前测量基于 Prestin蛋白的电致运动性的主要方法是一种间接的电生理技术,即膜片钳技术。该技术被广泛用于通过测量细胞所谓的非线性电容(NLC)来间接评估细胞电致运动性。但是,Song 等指出,在正常生理条件下,NLC的测量结果并不等同于运动性的直接测量。若要更令人信服地证实电致运动性确实属于压电效应,应采用传统的压电测量技术进行验证,尽管这在技术上存在挑战。此外,现有研究均集中在转染prestin的细胞中的电致运动性;若能直接对 prestin分子本身探测其压电性,将十分有意义。
4. 结论
从氨基酸到骨,压电效应在自然中的普遍存在暗示着它可能是生物学的一个普遍特征。这一想法与压电效应在生物系统中的生理意义通常并行。这个观点颇具吸引力,特别是当人们考虑到从深入理解该主题中可能产生的生物医学技术应用时。然而,目前尚缺乏系统的理解。在分子层面针对生物压电性机制的实验研究可能有助于实现这些应用。

















 29
29

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









