



2 生物传感器及其在传染病诊断中的应用
1 引言
在过去几年中,生物传感器因其在各种传染病诊断中的准确性、快速性、高灵敏度和高选择性,且成本低廉,而受到广泛关注 [1]。继 L.C. 克拉克发明首个用于氧气检测的真正氧电极传感器后 [2],现代生物传感器在灵敏度、选择性和多重检测空间方面已取得众多进展。通常,生物传感器可被描述为一种紧凑型分析装置,包含一个生物或生物来源的传感元件,并与物理化学换能器内部整合或紧密相连。生物传感器的两个基本工作原理包括生物识别和传感 [3]。生物传感器的主要组成部分包括生物识别系统(生物受体)、换能器和信号处理系统 [4]。尽管近年来在医疗保健设备和诊断技术方面取得了显著进展,但多种传染病的检测仍是一项重大挑战 [6]。目前广泛使用的基于实验室的传统病原体检测方法包括核酸扩增、培养、免疫测定和显微镜检查。
然而,这些实验技术大多存在根本性缺陷。例如,显微镜检查在许多临床发展阶段缺乏灵敏度,而培养过程耗时较长。同样,免疫测定虽然高度敏感,但劳动强度大且难以实现多重检测。此外,如聚合酶链式反应(PCR)等核酸扩增检测(NAATs)虽具有分子特异性,但样本制备方法复杂,且易出现假阳性 [5]。虫媒疾病对人类健康构成严重威胁,在全球范围内,其中疟疾最为严重。疟疾是一种通过蚊子传播给人类的致命性传染病,主要发生在热带国家 [7]。根据世界疟疾报告 2018 年 11 月发布的报告,2017 年疟疾病例达 2.19 亿例,高于 2016 年的 2.17 亿例。2017 年疟疾死亡人数估计为 43.5 万,这一数字令人担忧 [8]。
利用各种生物信息学工具和方法进行的计算研究有助于理解潜在的分子细节,这无疑将促进用于检测传染病的生物传感器及其他生物医学诊断技术的发展。通过分子建模和对接技术在原子尺度上全面了解结合机制,是生物传感器设计的必要前期工作。已有报道称,血红蛋白降解/疟色素形成是疟原虫生命周期中的关键步骤,也是新型抗疟疾药物发现的有前景的靶点。本文采用计算机模拟方法,结合同源建模和对接技术,研究血红素与二氢叶酸还原酶‐胸苷酸合酶(PfDHFR‐TS)以及血红素解毒蛋白(PfHDP)的结合情况,这两种酶均为抗叶酸类抗疟药的关键验证靶标。所获得的 PfDHFR‐TS 和 PfHDP 酶的三维(3D)结构及其与血红素结合相互作用的深入原子级信息,将为设计实验框架提供功能洞察,以开发携带突变酶的最优生物传感器模型来对抗疟疾。
1.1 生物传感器的原理和组件
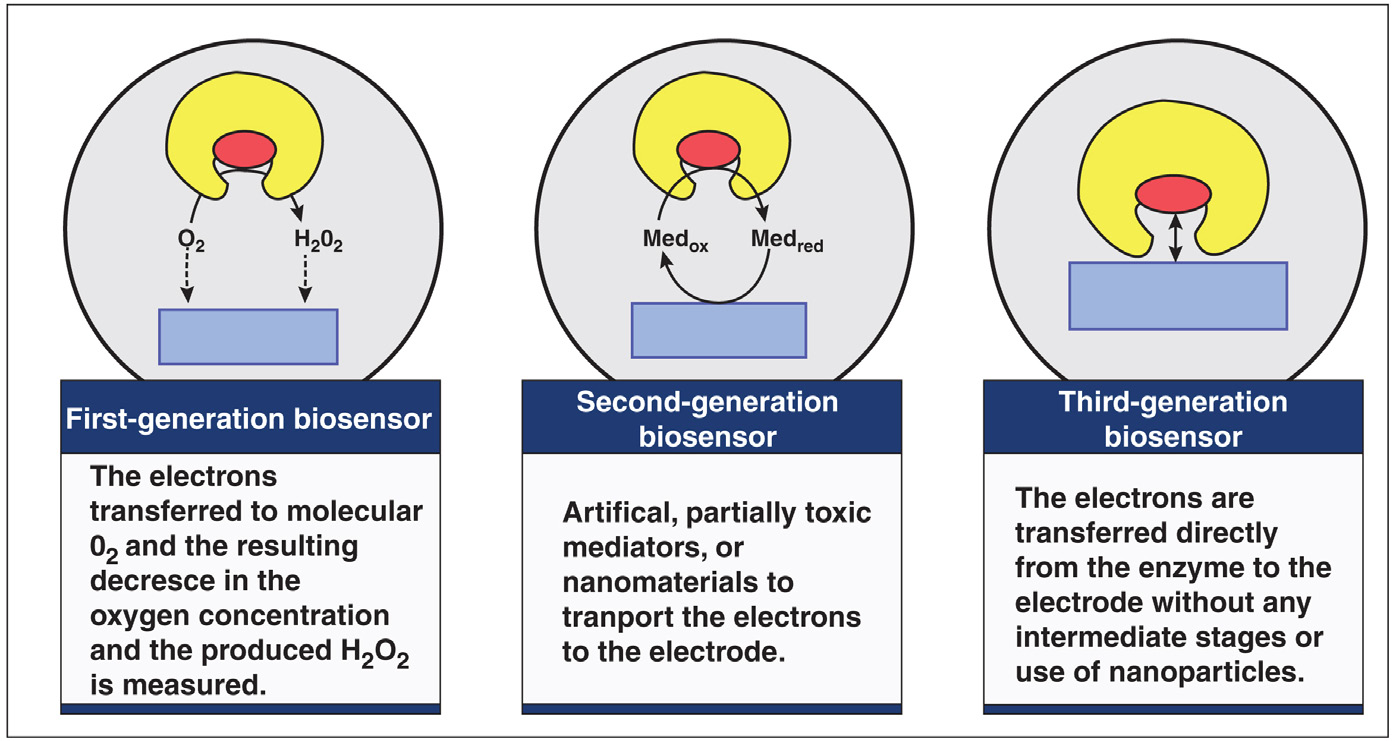
生物传感器设计的总体目标是能够在获取样本的即时检测点进行快速、方便的测试 [72]。如图 2.1 所示,生物传感器可分为三代。第一代生物传感器中,反应的正常产物扩散到换能器并引起电响应,而第二代生物传感器在反应与换能器之间引入特定的“介体”以产生更优的响应,第三代生物传感器则是反应本身直接引起响应,无需产物或介体的扩散参与。
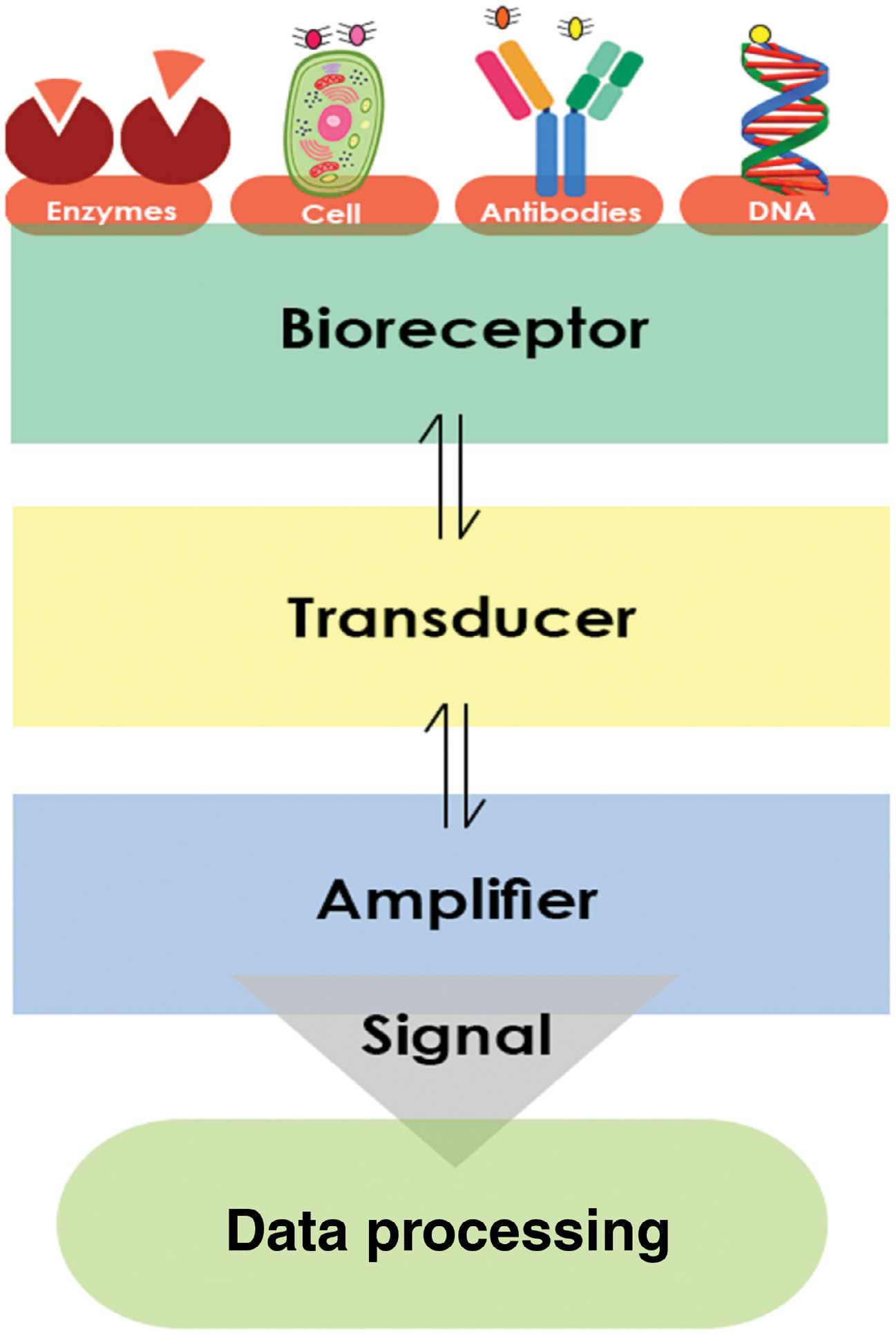
生物传感器可以定义为一种科学装置,包含固定的生物材料,如酶、抗体、核酸、激素、细胞器或完整细胞,这些材料能与分析物发生重要相互作用,并产生可测量的物理、化学或电信号。生物传感器的操作通常基于信号转导原理。生物传感器中广泛使用的三个主要组成部分如图 2.2 所示。
生物传感器的应用 [8] 被用于多种领域,如疾病监测、药物发现以及对体液(血液、尿液、唾液、汗液)中污染物、致病微生物和作为疾病指标的标志物的检测。图 2.3 显示了一个典型的生物传感器包含以下组成部分 [8]。
- 分析物 :分析物是一种需要测定其浓度的化合物(例如葡萄糖、尿素、药物、农药)。生物传感器基本上是通过将不同物质的生物作用转化为可测量信号,从而对其进行定量研究。例如,“富含组氨酸蛋白 II”是在用于检测疟疾的生物传感器中的“分析物” [9]。
- 生物受体 :它是一种能够特异性识别分析物的分子。酶、细胞、适配体、脱氧核糖核酸(DNA)和抗体在不同的生物传感器中作为生物受体。生物识别是指生物受体与分析物相互作用后以光、pH 值、质量变化或电荷形式产生信号的过程。目前开发的用于疟疾诊断的生物传感器中,常用的生物受体包括 DNA 适配体、HRP‐II 特异性抗体、针对疟原虫的生物素化 DNA 探针 [10,11,12]。
- 换能器 :在生物传感器中,换能器的功能是将生物识别事件转换为可测量的信号。这种能量转换方法称为信号化。大多数换能器产生光学或电信号,这些信号通常与分析物‐生物受体相互作用的数量成正比。金纳米粒子(GNPs)与聚二烯丙基二甲基氯化铵(PDDA)聚合物以及 GNPs 与聚(烯丙胺盐酸盐)(PAH)聚合物、电化学阻抗谱(EIS)电极以及用金纳米粒子功能化的丝网印刷金电极分别是适配体生物传感器、电化学生物传感器和免疫传感器中使用的一些换能器 [10,13,14]。
- 信号处理单元 :信号处理单元包含生物传感器的电子元件,用于分析转换后的信号并将其准备为可供显示的形式。它由复杂的电子电路组成,执行信号调理功能,如对信号进行放大以及将模拟信号转换为数字形式。处理后的信号随后由生物传感器的显示单元进行量化。显示单元由硬件和软件组合而成,以用户友好的方式呈现生物传感器的输出 [73]。输出信号可以是数值型、图形、表格或图像,具体形式取决于最终用户的需求 [74]。例如,用于疟疾诊断的生物传感器的输出可以通过比色法检测、阻抗响应、光学测量、电化学信号或荧光来检测 [15,16,17,18]。

1.2 生物传感器的分类
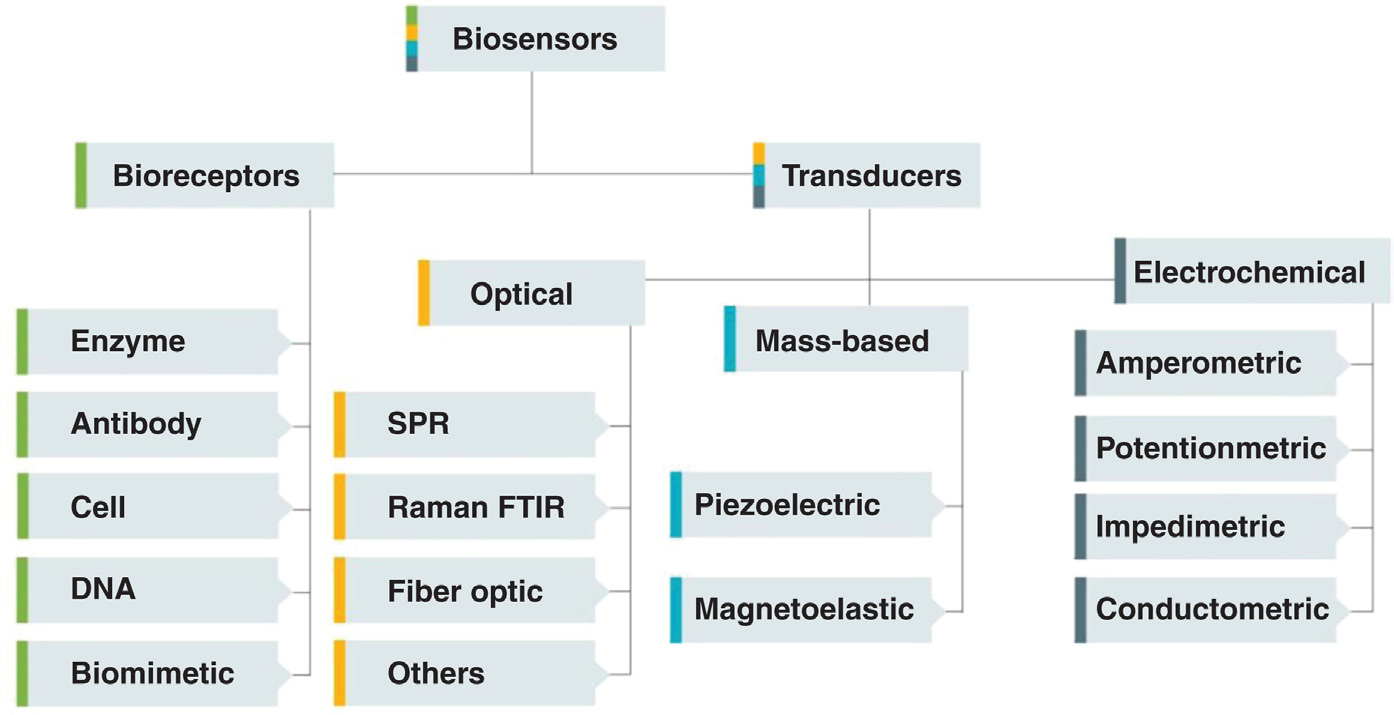
生物传感器可以根据其换能器类型、生物组分的性质或生物受体进行分类 [3,19],如图 2.4 所示。
生物受体构成传感器对目标分析物的识别系统。它基于通过生化机制识别分析物的分子种类。它们参与将感兴趣的分析物结合到传感器表面以进行测量。该受体可以是酶、抗体/抗原、核酸/DNA、细胞结构或仿生材料 [3]。
- 酶 :由于具有高效结合能力和催化活性,常被用作生物受体。厄普代克和希克斯于 1967 年报道了首个基于酶的生物传感器 [20]。此外,由于酶是天然蛋白质,在化学反应中不会被消耗,因此被选为传感器的潜在生物受体。
- 免疫传感器 :以抗体作为生物受体,可高亲和力地检测特定抗原 [3]。
- DNA 传感器 :通常基于单链核酸与互补链结合形成稳定氢键的特性而设计 [21,22]。首个细胞型传感器由迪维斯设计 [23]。
换能器是生物传感器的一部分,可将生物识别信号事件转换为可检测信号;后者可以是电化学的(电位法、电导法、阻抗法、安培法、伏安法)、光学的(比色法、荧光、发光、干涉测量法)、量热法(热敏电阻)、质量变化(压电/声波)或磁性 [3]。
- 电化学生物传感器 :通过将生物事件直接转换为电信号来分析生物样本的内容。首次报道的电化学阻抗谱出现在 1975 [24]。
- 光学生物传感器 :基于光电技术,其转导过程会因生物识别过程引起的物理或化学变化而导致入射光的相位、振幅、偏振或频率发生变化 [3]。
- 磁性生物传感器 :利用磁阻效应在微流控通道中检测磁性微纳米粒子,在灵敏度和尺寸方面表现出很高的效率 [25]。
1.3 当前常规实验室技术的比较 vis-a-vis 生物传感器在疾病诊断中的应用
及时准确的诊断和启动靶向抗菌治疗对于传染病的有效临床管理至关重要 [26,27,28,29,30,71]。特别是,由于持续暴露于致命病原体,虫媒疾病已成为热带和发达国家的巨大威胁 [31]。此外,在许多传染病中,早期症状通常是非特异性的,即在临床上无法与其他感染区分开来,因此需要明确的实验室信息。
在地方病情况下,分离感染性微生物作为一线方法是极为不可取的,但尽管如此,它仍是医学诊断中最直接和决定性的方法。目前有许多常规技术被用于医学诊断;然而,这些方法中的每一种仍然存在重大局限性。例如,尽管通过血清学技术进行病原体的分离和直接检测较为简单,但仍可能需要数天时间才能产生足够数量的抗体。同样,基于 PCR 的过程特别复杂,容易受到污染,并且需要熟练的人力以及笨重设备用于现场应用;此外,样本需要在极低温度下保存—这在农村环境中极具挑战性 [32]。类似地,微阵列技术成本高昂,难以在短时间内发挥作用并适用于即时诊断。在此背景下,生物传感器反而提供了一个易于使用、具有成本效益且灵敏的平台,能够快速识别病原体并预见有效的治疗方案 [26,27]。
此外,与生物测定或传统生物分析系统不同,生物传感器不需要添加试剂等额外的处理方法,并且其检测设计在设备构建中被永久固定 [33]。生物传感器在传染病检测中的应用非常广泛。可用于血液中病原体的早期检测 [34]、疫苗中毒力的检测、环境中危险病原体的检测,或用于治疗败血性伤口的类似绷带的设备 [35]。总之,与传统的生物亲和力测定相比,生物传感器具有更简便、快速和低成本的优势,同时具备高灵敏度和特异性。不同传染病的常见传统诊断方法在表 2.1 [5] 中进行了讨论。
1.4 恶性疟原虫二氢叶酸还原酶‐胸苷酸合酶和 PfHDP 与血红素相互作用的计算研究:疟疾的潜在生物标志物靶点
在各种虫媒疾病中,疟疾是一种常见的严重传染病。它在一百多个国家中构成了主要的公共卫生问题 [50]。
1.4.1 疟疾诊断的当前状况
成本和效益问题阻碍了疟疾快速诊断测试(RDTs)在全球范围内的进一步普及。早期诊断疟疾对于防止疟疾广泛传播至关重要,因此可通过适当的治疗降低患者死亡率。
- 显微镜分析 :显微镜分析被广泛认为是疟疾诊断的“金标准”。显微镜分析的优点包括较短的周转时间、通过薄血涂片确定物种的能力,即快速计算样本中感染的红细胞百分比,经济性好、便于样本保存以及简单的实验室基础设施。近年来报道了多种先进的显微镜技术,如二次散斑传感显微镜和荧光原位杂交技术,这些技术可提高疟疾诊断的检测概率 [60,61]。
- 基于抗体的 RDTs :RDTs 基本上是基于抗体的免疫层析试剂盒,可检测疟疾物种或疟疾抗原。RDT 的优点包括现场分析、无需电力即可运行、可检测多种物种、结果快速、经济性好、可由非专业或低技能技术人员操作,且操作和解释简便。
- 核酸扩增试验(NAATs) :NAATs 依赖于检测疟原虫核酸的技术。NAATs 特别推荐用于流行病学和研究研究。NAATs 的主要优势包括高效的灵敏度,能够检测每微升血液样本中一个寄生虫的能力,兼具定性和定量检测功能,有效检测多种寄生虫感染,能够同时处理多个样本(高通量处理),以及检测耐药菌株的能力。
但所有诊断技术都存在以下缺点:
1.
显微镜分析
:这种离场实验室方法需要熟练进行血涂片分析的人员及其培训,且结果相对较慢 [62]。
2.
快速诊断检测(RDTs)
:对低寄生虫密度不敏感。
3.
核酸扩增检测(NAATs)
:缺点是耗时长、成本高,在低浓度下重复性差,并且需要良好的实验室设施和熟练的技术人员 [63]。
不同的 Plasmodium 物种,如 Plasmodium vivax、P. falciparum、P. ovale 和 P. malariae 主要引起人类疟疾。最致命的物种是 P. falciparum,它通过雌性 Anopheles 蚊子叮咬传播 [51,52]。它会导致器官衰竭,并在脑毛细血管中积聚,导致晚期昏迷。在上述物种中,P. vivax 和 P. ovale 在感染早期阶段会影响肝脏 [51,52]。因此,选择直接针对肝阶段的药物以彻底治愈疾病非常重要 [52]。当肝细胞破裂时,裂殖子会攻击红细胞。在红细胞内相期间,裂殖子以不同形式进入。随后形成子代裂殖子。这些新产生的个体负责攻击新的红细胞。因此,靶向药物应尽量减少疾病的感染 [52]。在广泛的药物发现领域中,PfDHFR‐TS 和 PfHDP 均是疟疾研究中广为人知的靶标 [50,53,54]。P. falciparum 蛋白质对人类的肝阶段和血液阶段均具有可作用性 [50]。通过计算方法识别疾病标志物并预测其效应,有望开发出用于疟疾诊断和治疗的个性化工具。鉴于此,本文聚焦于疟原虫的血液阶段,旨在提供计算层面的见解,这将有助于利用同源建模和分子对接方法开发针对 P. falciparum 的血红素传感生物标志物(PfDHFR‐TS 和 PfHDP)。同源建模是解析蛋白质三维结构最常用的计算方法。分子对接是一种用于计算受体‐配体复合物结合亲和力和相互作用能的计算方法。
2 材料与方法
2.1 PfHDP 蛋白三维结构的测定
PfHDP 蛋白的三级结构使用 Modeller v 9.21 程序 [58] 生成。在模板选择时,对 BLASTp 搜索进行了 PDB 数据库 [66] 除了 I‐TASSER 工具 [67] 用于寻找具有最高同一性和较低 E 值的 X 射线晶体结构。最终,选择 PDB ID: 5NV6 的晶体结构作为建模模板,其查询覆盖率为 27%,同一性为 77%。通过 Modeller 程序生成的 200 个不同模型中,根据最高的 DOPE(离散优化蛋白质能量)分数选出最佳模型,并进一步使用 GalaxySite [59] 程序进行环区和侧链优化。
2.2 恶性疟原虫二氢叶酸还原酶‐胸苷酸合酶与血红素的对接
使用 AutoDock Vina 软件对 PfDHFR‐TS(PDB ID: 3UM6)和血红素(PubChem ID: 26945)的晶体结构进行对接 [56]。设置网格盒参数以形成足够尺寸的空腔,从而容纳各化合物进入蛋白质的结合位点。采用拉马克遗传算法搜索最佳配体构象,其余参数均设为默认值。最终构象根据 AutoDock 评分函数进行聚类和排序。选择结合亲和力最低的对接构象,进一步使用 PyMOL 工具(http://www.pymol.org/)和 Discovery Studio Visualizer 4.0 软件分析结合界面及相互作用残基 [57]。
2.3 PfHDP 与血红素的对接
使用与 PfDHFR‐TS–血红素复合物相同的方案,将建模的 PfHDP‐TS 与血红素复合物进行对接。
2.4 结构验证建模的结构
所构建的 PfHDP 模型通过多种结构验证工具进行了立体化学验证,包括 PROCHECK [68], ProSA [55], VERIFY3D [69], 和 ERRAT [70] 网络服务器。
所有这些结构验证工具采用不同的方法计算定量评分,可用于评估模型质量,并指导进一步筛选最精确的蛋白质结构。PROCHECK 通过评估拉马钱德兰图中二面角(φ 和 ψ)的准确性来验证蛋白质结构的立体化学质量。ProSA 计算一个整体 Z‐分数,可用于评估计算所得蛋白质模型相对于 PDB 中已知晶体结构和 NMR 结构的结构质量。VERIFY3D 用于分析 3D 蛋白质模型与其自身氨基酸序列的兼容性。ERRAT 输出一个整体模型质量因子,用于检验非键相互作用的原子间作用。VERIFY3D 和 ERRAT 得分越高,表明结构越稳定可靠。
表 2.2 总结了本项生物信息学研究中所使用的各种计算工具和数据库。
| 工具和数据库 | 描述 | 参考文献 |
|---|---|---|
| BLAST | 查找序列之间的局部相似性区域 | [65] |
| 蛋白质数据库 | 存储大型生物分子三维结构的数据库,包括蛋白质和核酸的分子核酸 | [66] |
| Modeller 9.19 | 蛋白质三维结构的同源或比较建模 | [58] |
| VERIFY3D | 确定原子的相容性,具有自身氨基酸的模型(3D),通过分配一个结构类别来确定序列(1D)基于其位置和环境 | [69] |
| ERRAT | 分析非键相互作用的统计信息,不同原子类型之间的相互作用,并绘制误差函数的值,相对于 9 残基滑动窗口的位置,通过与高度精修结构的统计信息进行比较计算得出 | [70] |
| PROCHECK | 通过分析残基的几何结构和整体结构几何来检查蛋白质结构的几何形状 | [68] |
| ProSA | 计算蛋白质结构的总体质量评分。特定输入结构。如果该分数超出天然蛋白质特征范围的结构可能包含错误 | [55] |
| AutoDock Vina | 蛋白质‐配体对接 | [56] |
| PyMOL | 生物分子的分子可视化结构 | [64] |
| Discovery Studio 4.0 | 用于分析和可视化大分子的综合软件套件 | [57] |
| ## 3 结果与讨论 |
3.1 血红素与 PfHDP 及 PfDHFR‐TS 的结合
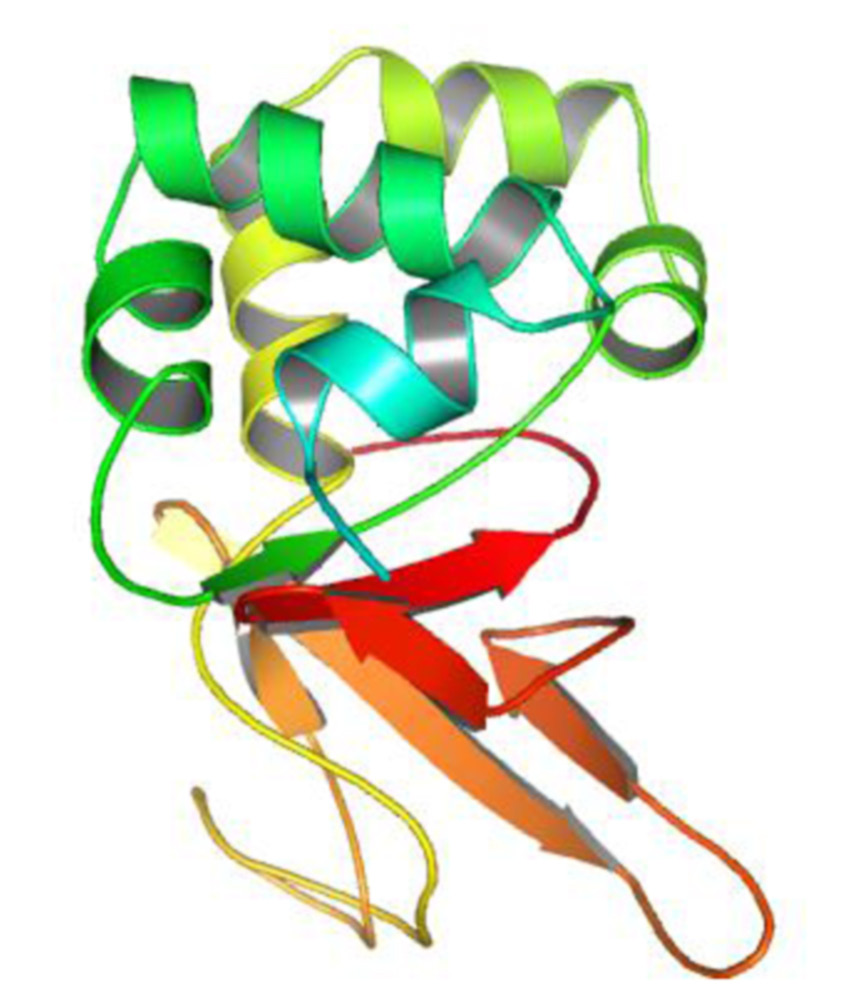
使用 Modeller 软件预测了 PfHDP 的三维结构。建模的 PfHDP 蛋白(图 2.5)包含六条反平行的 β‐折叠片和五个 α‐螺旋。
使用多种结构验证工具进一步评估了预测的 3D 模型的质量。PfHDP 的 PROCHECK 结果(图 2.6A)表明所构建的结构具有可靠的立体化学质量,拉马钱德兰图中超过 73% 的残基位于允许区域。为了检测结构中的潜在错误,Z‐score 通过 ProSA‐web 进行计算。
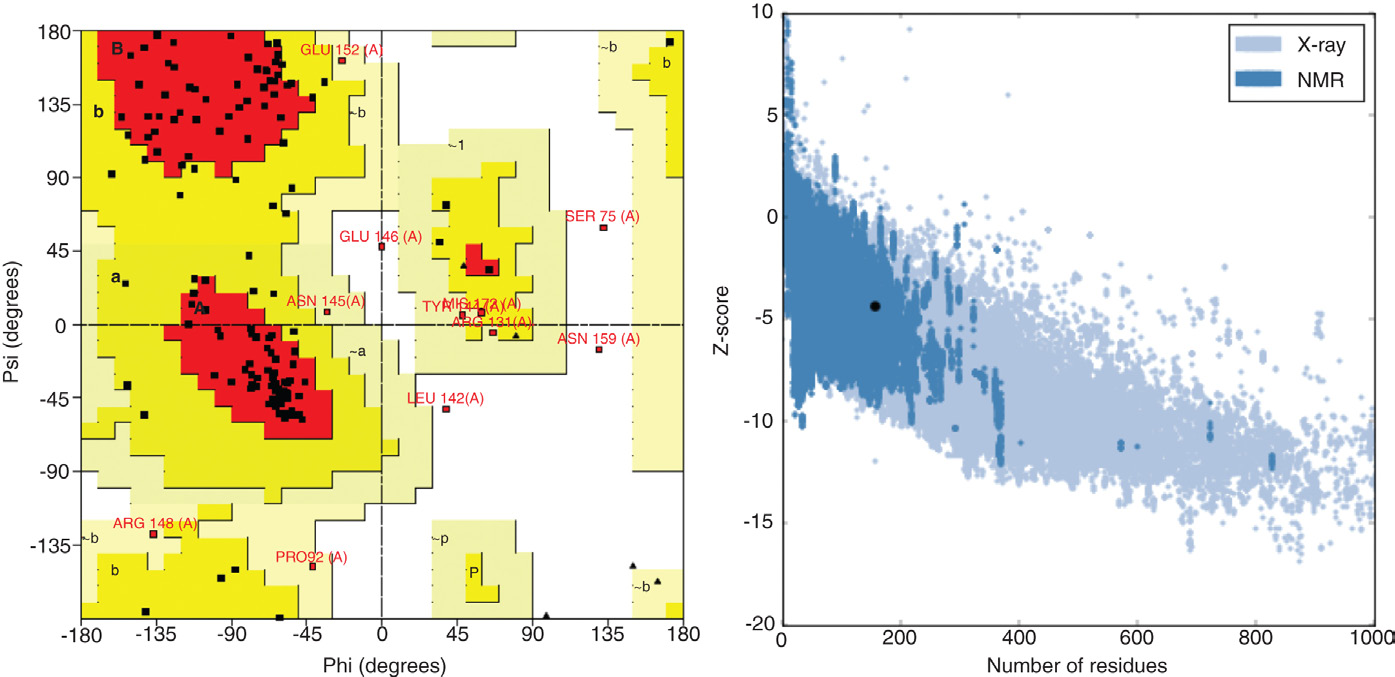
 使用PROCHECK服务器生成的PfHDP模型的拉马钱德兰图。(B) ProSA‐web Z‐scores of all protein chains in PDB determined by X‐ray crystallography (lightblue) 和核磁共振波谱 (dark blue) 相对于其长度的结果)
使用PROCHECK服务器生成的PfHDP模型的拉马钱德兰图。(B) ProSA‐web Z‐scores of all protein chains in PDB determined by X‐ray crystallography (lightblue) 和核磁共振波谱 (dark blue) 相对于其长度的结果)
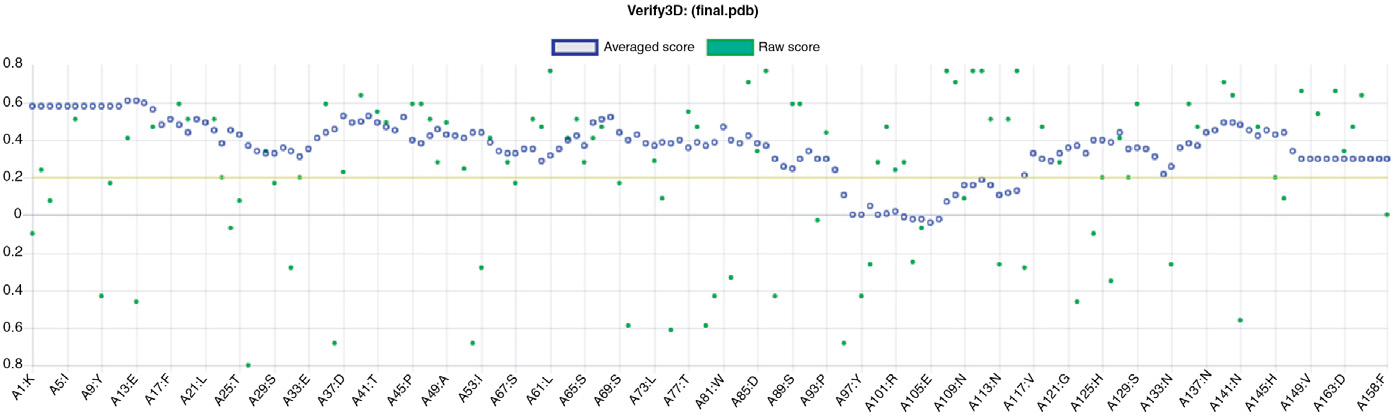
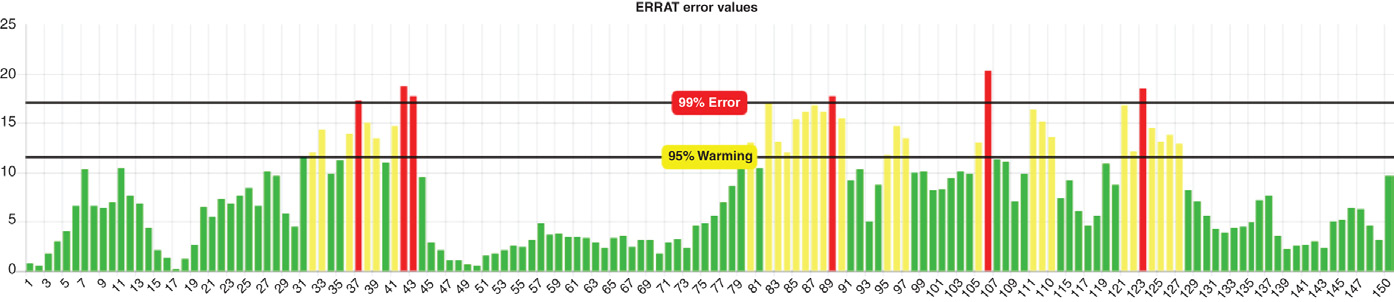
程序。超出天然蛋白质特征范围的得分表明结构有误。在本研究中,Z‐score 值显示 PfHDP 落在 PDB 中实验测定的蛋白质的范围内(图 2.6B)。此外,使用其他多种结构验证工具获得的高分值,即 VERIFY3D 和 ERRAT 展示了所建模结构的整体准确性(图 2.7 和 2.8)。
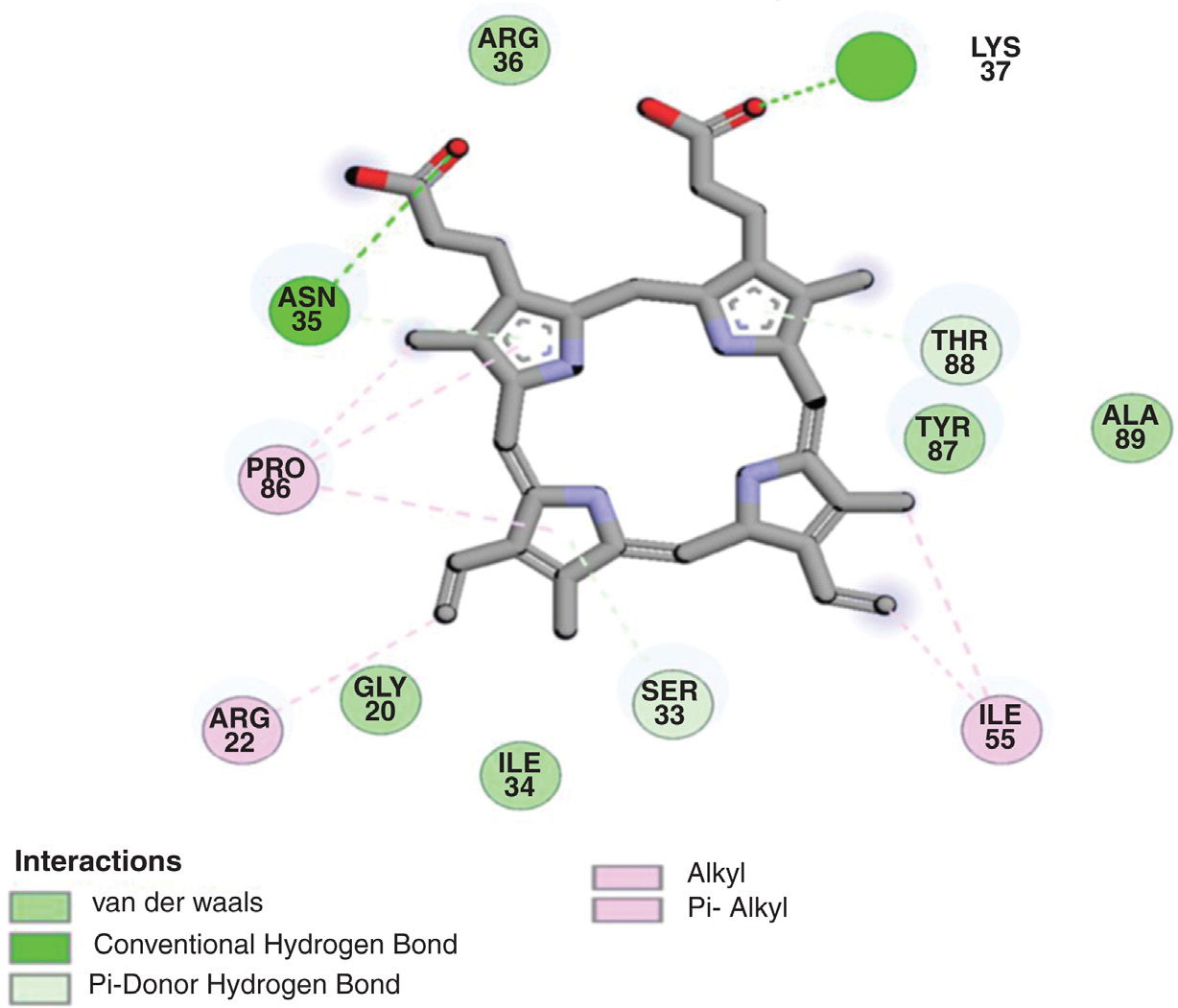
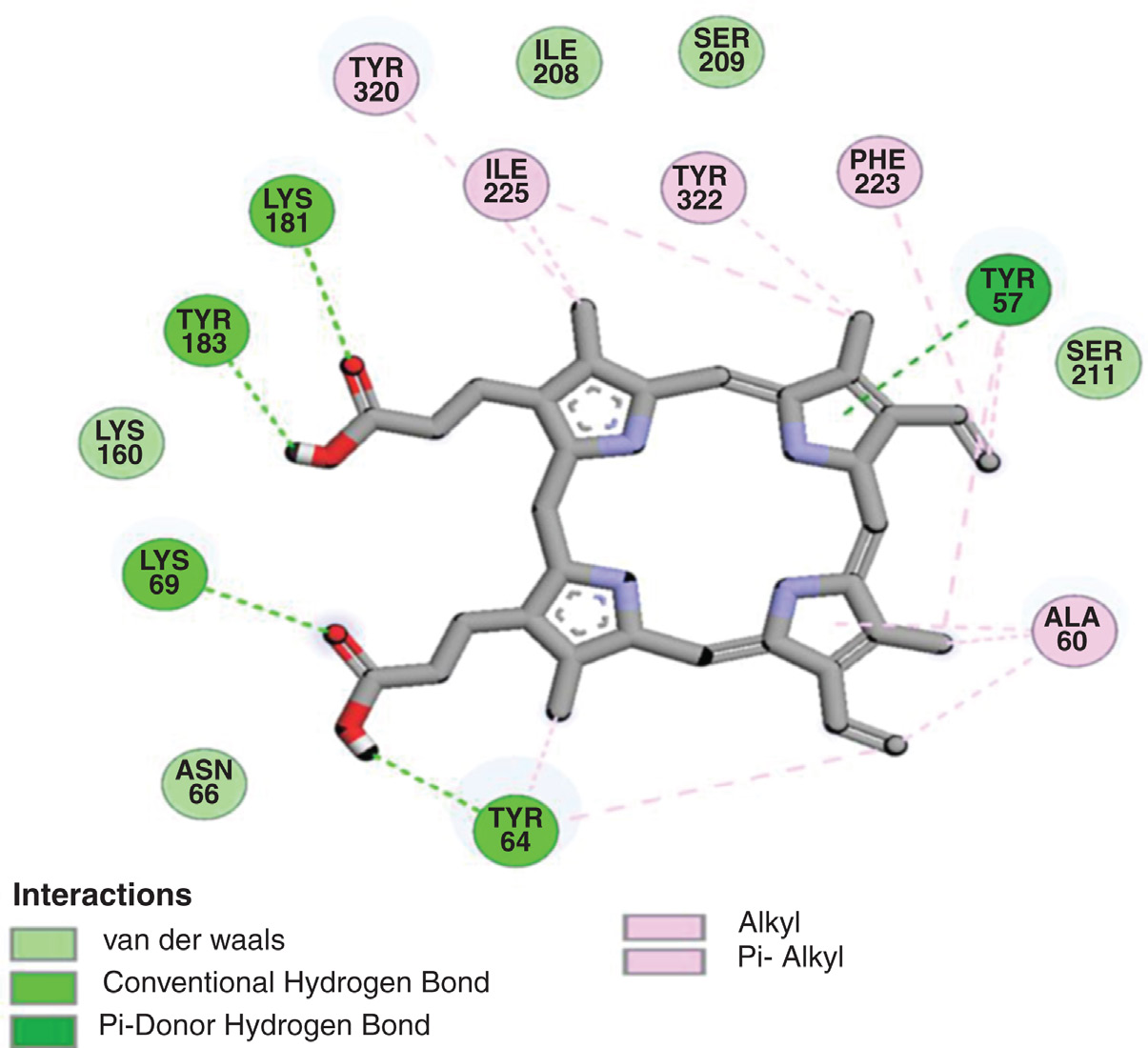
结构测定后,进行了 PfHDP 和 PfDHFR 与血红素的分子对接,以探索它们的潜在结合界面。PfHDP–血红素和 PfDHFR–血红素对接复合物的结合相互作用如图 2.9 和 2.10 以及表 2.3 所示。PfHDP–血红素复合物的对接三维结构显示,PfHDP 的四个残基 ASN35、LYS37、SER33 和 THR88 通过四个氢键(H‐bonds)与血红素蛋白发生相互作用。相互作用的氢键距离在 2.723–3.943 埃范围内。PfDHFR–血红素复合物的对接三维结构显示,PfDHFR 的五个残基 LYK69、LYS181、TYR183、TYR64 和 TYR57 通过五个氢键(H‐bonds)与血红素蛋白发生相互作用。相互作用的氢键距离在 2.065–4.107 埃范围内。
4 结论
最近,为开发更有效且经济实惠的药物和预防性疫苗以彻底根除疟疾,已启动多项新举措。为此,诺华和葛兰素史克等多家制药公司报告称,在筛选约 200 万种小分子的化学文库以寻找抗疟疾特性后,发现了超过 20,000 个“高可成药性”的初始命中分子。此外,目前学术界的研究重点集中在阐明特定通路靶点。其中一项高通量筛选工作聚焦于疟原虫特有的血红素解毒途径——疟色素形成。本章讨论了生物传感器在诊断疟疾等传染性疾病以及发病机制疾病中的重要性。
通过计算方法识别疾病标志物并预测其效应,有望为疟疾的诊断和治疗开发个性化工具。先前的研究表明,PfDHFR‐TS 和 PfHDP 均为疟疾有前景的生物标志物靶点。基于此背景,本研究采用计算方法深入分析了 PfHDP 和 PfDHFR 与血红素的分子相互作用。对接复合物显示,PfHDP–血红素的结合能为 −7.47 kcal/mol,PfDHFR–血红素的结合能为 −9.61 kcal/mol。对接形成的 PfHDP 与血红素复合物以及 PfDHFR‐TS 与血红素复合物的结合能分别为 −7.47 kcal/mol 和 −9.61 kcal/mol,表明血红素对 PfDHFR‐TS 的传感强度优于对 PfHDP 的传感强度。
本文所讨论的生物传感器现状以及相关计算研究,将为传染病治疗干预措施的设计提供有益见解。未来必须开发新的方法,以验证这些命中化合物在寄生虫内的作用机制。
| 参数 | PfHDP | PfDHFR |
|---|---|---|
| 结合能 (kcal/mol) | −7.47 | −9.61 |
| 键类型 | 氢键 | 氢键 |
| 结合残基和原子 |
ASN35–O
LYS37–O SER33–π轨道 ASN35–π轨道 THR88–π轨道 |
LYS69–O
LYS181–O TYR183–H TYR64–H TYR57–π轨道 |
| 抑制常数 (μM) | 3.34 | 89.61 |
| 分子间能量 (kcal/mol) | −10.45 | −12.6 |
| 静电能 (kcal/mol) | −3.43 | −4.85 |

















 385
385

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









