



爬行动物中的性别反转:生殖异常还是推动进化的强大力量?
摘要
性别是由基因、环境还是两者共同决定的?在这篇综述中,我们描述了爬行动物中性别决定机制的多样性,重点关注表现出基因‐环境相互作用的系统。我们总结了来自野外和实验室的证据,证明爬行动物中存在环境性别反转,并探讨这是否是一种广泛存在的进化机制,影响脊椎动物性染色体的进化和物种形成。性别决定系统存在于遗传因素和环境因素影响的连续谱系中,模糊了过去被认为是遗传型性别决定与温度依赖型性别决定之间的严格二分界限。在此连续谱系中,我们发现具有明显分化的异形性染色体的物种(Pogona vitticeps、Bassiana duperreyi、Eremias multiocellata, Gekko japonicus)、具有弱分化同形性染色体的物种(Niveoscincus ocellatus)以及仅具有微弱可遗传倾向决定性别的物种(Emys orbicularis, Trachemys scripta)均存在性别逆转的可能性。我们认为,性别逆转是推动爬行动物性别决定机制可塑性和多样性产生与维持的重要进化力量。
在脊椎动物中,性腺结构具有广泛的共通性
在脊椎动物中,性腺结构具有广泛的共通性 [Morrish 和 Sinclair,2002;Rhen 和 Schro‐e der,2010],因此调控性腺分化的基因和调控过程在脊椎动物之间也具有高度的共通性也就不足为奇了 [萨尔等人,2004;Cutting 等人,2013]。然而,对于调控性别决定、引导受精卵走向雄性或雌性发育轨迹的基因和调控过程而言,情况却并非如此。脊椎动物物种间性别决定机制的变异确实非常显著,而爬行动物极大地贡献了这种多样性 [Bachtrog 等人,2014]。因此,爬行动物的性别为研究提供了丰富的领域,有助于更广泛地理解脊椎动物性别决定 [加姆布尔等人,2015;霍利等人,2015;彭内尔等人,2015]。
许多爬行动物具有遗传性别的决定(GSD)系统。其中,许多龟类和蜥蜴具有雄性或雌性异配性(ZW和 XY,某些蜥蜴中为XXY),而蛇类则具有雌性异配性 (ZW、ZZW或ZWW)[Solari, 1993;Olmo, 2005; Ezaz 等人,2010;Georges 等人,2010] (图1)。爬行动物也是唯一存在专性孤雌生殖的羊膜动物,这种现象通常起源于种间杂交[Neaves 和 Baumann,2011]以及一些兼性孤雌生殖的显著例子[Booth 等人,2012]。许多其他具有遗传性别的决定(GSD)的爬行动物物种拥有同态性染色体,在细胞学上难以区分[加内什和拉曼,1995;Ezaz 等人,2010]。另一些爬行动物则具有温度依赖型性别决定(TSD),其性别由卵孵化时的温度决定,几乎没有直接的遗传影响证据[Bull, 1980; Ewert 等人,1994;Harlow,2004]。最近研究表明,这两种看似独立的性别决定方式(GSD和TSD)在某些物种中会相互作用,导致染色体性别发生逆转[Quinn 等人,2007;Radder等人,2008年;Holleley 等人,2015]。本文综述了爬行动物中的性别逆转现象,其在野外的普遍性,对性别决定机制整体理解的可能影响,以及其进化意义。
什么是性别逆转?环境机制与突变机制的对比
性别逆转发生在性染色体正常的遗传影响下本应发育为某一性别的个体,由于环境或突变的影响而转向发育为另一性别。这种对调控性腺表型过程的干扰可能导致可存活但相反的性表型,或导致性腺发育不良和适应度受损。本文综述仅限于在实验室条件或自然环境中产生可存活性表型的性别逆转情况。性反转个体被称为不一致个体,因为其表型性别与其基因型性别相反。这种不一致可朝任一方向发生:个体可能是遗传学雄性但表现为解剖和功能上的雌性,或是遗传学雌性但表现为解剖和功能上的雄性。
明确检测性别逆转的关键在于拥有特征明确的性染色体系统,以及关于性染色体序列和基因含量的基因组信息。爬行动物中同态性染色体的广泛存在可能掩盖了该类群中性别逆转的发生。多年来,充分区分性染色体的能力仅限于相对粗糙的G带和C带技术,或依赖于大小或粗略形态的差异。当性染色体为微小染色体或形态差异较细微时,这些性染色体很可能是隐性的,因而容易被忽视 [Ezaz et al., 2009, 2010]。更灵敏的细胞遗传学技术,如比较基因组杂交,已在多种物种中揭示出此前未被发现的隐性性染色体[Traut et al., 1999],包括性微小染色体以及近期进化而来且形态分化程度较低的性染色体[Traut et al.,1999; Ezaz et al.,2005;Kawai et al.,2007]。使用灵敏的分子细胞遗传学方法非常重要,因为异态元件(Y或W)上高度重复的重复序列使其通过分子方法进行测序和组装尤为困难[Tomaszkiewicz et al.,2016]。
性别逆转的一种途径是性别决定相关基因(例如哺乳动物中的主控性别决定基因SRY,以及许多其他后生动物中的DMRT1或其旁系同源基因)或其相关调控区域发生突变。然而,需要认识到的是,只要这种干扰不损害功能性性别表型,性别逆转也可能由主控性别决定基因下游的调控基因及其产物在表达、传递或接收方面的改变所引起。一个很好的例子是芳香化酶(CYP19A1),它在爬行动物中不太可能是主控性别决定基因,但当其受到抑制时,会导致雌性向功能性雄性表型的转变[Wibbels 和 Crews, 1994;Belaid 等, 2001;Britt 等, 2001; Lance, 2009]。因此,任何损害芳香化酶功能的突变都可能引发性别逆转,而不会影响个体的存活能力。
绝大多数性染色体研究集中在恒温脊椎动物,即哺乳动物和鸟类。这些物种的性染色体相对稳定且高度分化,外部环境对性别决定几乎没有或很少产生影响。这导致了一种较为偏颇的观点,即性别逆转仅通过突变过程作为遗传异常发生[Eggers et al., 2014; Ohnesorg et al., 2014]。然而,性别逆转既可由基因突变引起,也可由环境对性别决定和/或性别分化级联中基因的影响所致,后者是本综述的重点。新兴理论和越来越多的实证证据表明,环境诱导的性别逆转是一种普遍且自然发生的现象,可能显著而迅速地影响物种进化,尤其是在鱼类 [Chan和Yeung,1983;Shao等,2014;McNair Senior等,2015]中,几乎可以肯定也在两栖动物 [杜农等,1990年;Miura,1994;Alho 等,2010]中存在。性别逆转 increasingly 被认为是导致外温动物性染色体不稳定性的 一种机制 [Ogata等,2003;Perrin,2009;Holleley等,2015]。理解性别逆转的动态对于理解爬行动物进化以及预测气候变化体制对爬行类多样性未来影响至关重要。
野生爬行动物种群中环境性别逆转的确诊案例
激素操控和外源性化学物质暴露可导致具有性染色体的两栖动物发生性别逆转[杜农等,1990年; Orton 和 Tyler,2015],类似的操作也已在温度依赖型性别决定(TSD)爬行动物中进行[Bull 等,1988; Belaid 等,2001;Warner 和 Shine,2008; Mizoguchi 和 Valenzuela,2016] ,使其性别与孵化温度所预期的性别相反。然而,这些研究对于野外实际情况的相关性仍存在一些推测。
关于影响爬行动物性别决定的自然发生的基因‐环境相互作用,目前可用的信息非常有限。在具有已知性染色体的爬行动物中,仅在两个进化上独立的卵生飞蜥科和石龙子科蜥蜴实例中观察到自然发生的性别逆转现象。
对爬行动物性别逆转研究最深入的例子是中部鬃蜥 (Pogona vitticeps)。该物种具有可识别的ZZ/ZW微型性染色体 [Ezaz 等,2005] ,在大多数温度下子代性别比例均衡 [Quinn 等,2007;Holleley 等,2015]。然而,在高温孵化条件(>32 °C)下,ZZ(通常为雄性)性染色体的信号被抑制,导致性别比例逐渐偏向雌性 [Quinn 等,2007;Holleley 等,2015]。分子和细胞遗传学技术已明确证实,这种雌性偏态是由性别逆转(即雌性化的ZZ个体)引起的,而非由性别特异性的子代死亡率所致 [Quinn 等,2007;Holleley 等,2015]。最重要的是,性别逆转的 P. vitticeps 在野生种群中自然存在,并且具有生殖能力 [Holleley 等,2015]。
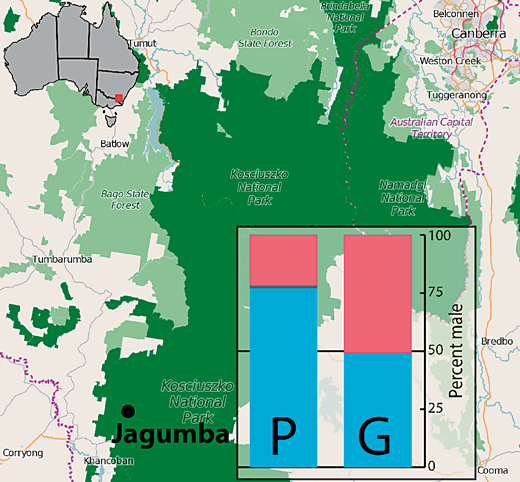
在第二种情况下,东部三线石龙子(Bassiana duperreyi)的环境条件与染色体性别决定共同作用以决定性别[Shine等人,2002年;Radder等人,2008年; Quinn等人,2009]。杜氏滑趾壁虎具有雄性异配的性别决定系统,其中Y染色体大部分为异染色质,且明显小于X染色体[Donellan,1985年;Matsubara等人,2016]。在实验室中,染色体为雌性(XX)的个体在低卵孵化温度下会发生雄性化[Radder等人,2008年; Quinn等人,2009]。正在进行的研究已发现,在杜氏滑趾壁虎高山种群的巢穴中存在高比例的性别逆转现象(28% XX雄性)(图2),但目前尚不清楚该物种的性反转个体是否具有生殖能力。
这两个案例表明,在自然界中,具有分化性染色体的爬行动物物种可能发生性别逆转。然而,这些例子仅占具有性染色体的爬行动物物种中极小的一部分,因此目前尚无法准确估计性别逆转的发生率。据我们所知,尚未有其他已发表的实例显示,对具有明确性染色体的爬行动物物种进行了严格的温度孵化实验且未观察到温度效应。关于温度对 GSD物种子代性别比例影响的发表数据缺乏,可能是因为此类实验通常未被报道,阴性结果被认为过于明显而不适合发表,或者当实验出于其他目的进行时,阳性结果被视为异常。我们认为,孵化实验偏向于无性染色体物种的现象,部分源于一种潜在的观点,即认为TSD和GSD是截然对立的状态而非相互作用的状态[Sarre et al.,2004]。
混合模型性别决定和推断的性别逆转
尽管上文讨论的P. vitticeps和杜氏滑趾壁虎是性别逆转现象最明确且研究最深入的爬行动物案例,但越来越多的证据表明,在基因与环境共同作用决定性别的混合模型系统中,这类情况十分普遍。虽然在这些物种中尚未明确证实性别逆转的存在,但实验和生态学证据表明性别逆转确实正在发生。
关于混合型或过渡性性别决定系统存在的最早证据之一来自龟类Emys orbicularis。尽管该物种具有表现出一种看似温度依赖型的性别决定模式 [Zaborski 等,1982, 1988],在性别决定的关键温度下可检测到遗传效应,而在该温度下温度的影响尚不明确 [Zaborski 等,1982, 1988;杜农等,1990; Girondot 等,1994]。在关键温度下,表型性别与 H‐Y抗原的表达一致。具体而言,在体细胞和性腺组织中,雄性为H‐Y阴性(H‐Y – ),雌性为H‐Y阳性( H‐Y + )。在其他温度下,H‐Y抗原在体细胞中的表达则不一致,即表型性别与H‐Y抗原表达解耦,推测是因为温度覆盖了遗传倾向。因此,似乎具有温度依赖型性别决定(TSD)的圆斑蜥蜴(E. orbicularis)其性别决定存在潜在的遗传成分,但在极端温度下被温度所覆盖。令人意外的是,当检查野外捕获动物时,发现表型性别与遗传性别不一致的情况较少(6%的表型雌性和 11%的表型雄性)[Girondot 等,1994]。自然巢穴中的热环境似乎更有利于遗传因素而非温度因素,同时也表明在实验室条件下以及在野外低频率地发生了圆斑蜥蜴(E. orbicularis)的性别逆转。要确切识别性别逆转,还需开发性连锁分子标记。
在最近描述的一个例子中,眼斑雪石龙子( Niveoscincus ocellatus),一种沿塔斯马尼亚海平面至高地地区1200米海拔和气候梯度分布的小型卵胎生蜥蜴,表现出遗传性别决定被温度覆盖的特征[Wapstra et al., 2004, 2009; Pen et al., 2010]。尽管该物种的性别决定系统目前尚不清楚,但来自不同海拔的种群对母体妊娠温度具有不同的响应[Pen et al., 2010]。高海拔种群无论妊娠温度如何,后代性别比例均等;而低海拔种群在较低孵化温度下则表现出雄性偏态性别比例。虽然尚未在眼斑雪石龙子中鉴定出异形性染色体,但其近缘物种具有XY 异配性别,且遗传性别决定是谱系中的祖先特征[Donellan, 1985]。采用比较基因组杂交或其他高分辨率细胞遗传学技术,可能揭示该物种中隐性性染色体的存在,并为证明其发生性别逆转提供依据。
在第三种情况下,胎生蜥蜴Eremias multiocellata的子代性别比例在低温下表现为雌性偏态(25 °C时雄性占15.4%),在高温下表现为雄性偏态(35 °C时雄性占83%)[Zhang et al., 2010]。这被解释为温度依赖型性别决定(TSD),但涉及性染色体染色体现已通过比较基因组杂交在该物种中被鉴定出来[Wang 等,2015],表明在实验室中温度正在覆盖潜在的性染色体系统。该物种中尚未发现性反转个体,但这些数据表明性别逆转很可能发生。
眼斑夜守宫和多眼斑壁虎中性别逆转的证据使我们注意到,热效应对性别决定的影响似乎不仅限于产卵物种,外温动物的胎生物种其性别决定也可能受到母体妊娠期间环境的影响。
卵生壁虎在性染色体上表现出显著的进化不稳定性,已发生多达25次独立的转变 [Gamble et al., 2015]。尽管已有明确证据表明存在频繁的性染色体更替,并且现已具备完善的分子工具包来研究性别逆转,但在壁虎中仍尚未确定性别逆转的确凿例证 [Gamble et al., 2015]。早期研究表明,在日本壁虎中可能存在异形性染色体与温度依赖型性别决定(TSD)的共现现象 [吉田和伊藤, 1974;Tokunaga, 1985]。然而,这些观察结果需要通过同步进行的孵化实验和核型分析加以验证,以排除两项独立研究可能无意中针对了不同的隐存种或常见的种间杂交种的可能性 [户田等人,2001,2006;萨尔等人, 2004;Gamble, 2010]。
性反转对适合度的影响
性反转研究主要集中在哺乳动物或鸟类,其中性别逆转源于基因表达的突变影响。这种分类学偏见导致了普遍态度认为性反转爬行动物本质上是有缺陷的,因此适合度较低[Pokorna和Kratochvil,2016]。当性别逆转及后续繁殖产生无法存活的WW或YY个体时,较低适应度是一个合理的假设。然而,在两种已有充分记录的爬行动物性别逆转案例中(P. vitticeps和杜氏滑趾壁虎),性别逆转仅发生在同配性别(ZZ或 XX)中,从而消除了因产生无法存活的WW或YY后代而导致的适合度亏损[Bull; Quinn等;Schwanz等]。
鱼类实验研究表明,环境性别逆转可能会损害某些与适应性相关的性状(如生长速率),但其他性状则不受影响(如存活率)[McNair Senior et al.,2012]。然而,我们认为所观察到的对生长速率的负面影响更可能是对用于实验性诱导性别逆转的内分泌干扰化学物的一种暂时性反应[McNair Senior 等 2012]。
尽管可比的爬行动物研究较少,但初步数据显示,性别逆转并不像先前假设的那样有害。事实上,ZZ型性别逆转雌性鬃狮蜥所产生的卵数量几乎是ZW一致型雌性的两倍[Holleley 等2015]。这表明性别逆转可能带来适合度上的益处。
我们还知道,性别逆转可能对影响适合度的行为特征产生显著影响[Gatewood 等,2006; Kopsida 等,2013;Li 等,2016;Saunders 等, 2016]。例如,发生性别逆转的鬃狮蜥表现出一系列独特的雄性和雌性形态及行为特征,使其能够对选择压力作出差异化的响应[Li 等,2016]。最重要的是,在测试新奇恐惧和领地探索时,性别逆转的鬃狮蜥明显比遗传一致的雄性和雌性个体更具大胆性[Li 等, 2016]。大胆性是一种与适合度相关的重要行为表型,因为它会影响个体寻找和获得交配机会的能力[Mazué 等,2015;Sih 等,2015]。然而,在捕食者丰富的环境中,更大胆性会带来适合度上的权衡,因为更显眼的行为也可能增加被捕食导致的死亡风险[Sih 等,2015]。
在实验室研究中观察到的性反转个体具有高繁殖力和增强的大胆性,但这是否能在野外条件下转化为适应度提升仍有待验证。然而,显然不能假设爬行动物或其他脊椎动物中的性反转个体具有较低适应度[Saunders et al.,2014, 2016]。事实上,情况可能恰恰相反。在野外条件下,对自然发生的性反转个体的终生适应度进行表征,是理解该过程在爬行动物中进化动态的下一个关键步骤。

性别逆转维持性别决定模式的可塑性和多样性
最近在野生爬行动物种群中发现的性别逆转现象表明,环境性别反转如何促使温度依赖型性别决定(TSD)迅速进化[Holleley 等2015]。这一发现与普遍认为性染色体进化是单向“陷阱”的观点相悖。根据该假说,温度依赖型性别决定(TSD)是祖先状态,物种可能保持在此状态,或者随后转变为基因型性别决定(GSD)模式。获得遗传性别决定位点。通过倒位抑制重组并导致性染色体分化,从而阻止了返回到温度依赖型性别决定 (TSD)状态的可能性 [Bull 和 Charnov,1977; Bull,1983;Pokorna 和 Kratochvil,2009; Bachtrog 等,2014]。尽管这种性染色体进化的“陷阱”模型在哺乳动物中具有说服力,但在其他脊椎动物中,性别决定似乎更加可变和易变。事实上,获得一个遗传性别决定位点并不一定意味着其所在染色体的命运已被锁定 [Wright 等,2016]。新型性别决定模式的进化存在多种可能路径 [Quinn 等,2011; Sarre 等,2011;Schwanz 等,2013]。实证研究 [ Holleley 等,2015]和理论研究 [Quinn 等,2011]表明,仅通过改变雄性或雌性决定因子基因表达的阈值,即可轻松获得所有可能的性别决定模式(XY、ZW、TSD)。在不确定且易变的环境中,可变的性别决定机制可能具有优势,而性染色体更替过程可能导致参与性别决定的基因多样性增加。这一过程或许可以解释为何目前尚未在爬行动物中鉴定出主控性别决定基因。性别决定位点的反复且独立的进化捕获使得在不同爬行动物种系甚至亲缘关系非常接近的物种之间,很难有一个单一基因参与性别决定(图1)。
重申这一点,我们注意到目前已确定的3种爬行动物候选性别决定基因在有鳞类(NR5A1 P. vitticeps; Deakin 等 [2016])和龟鳖目(WT1 Glyptemys insculpta; Montiel 等 [2016]以及 CIRBP 蛇颈鳄龟; Schroeder 等 [2016])中各不相同。
爬行动物性别逆转与气候变化:未来会是一个没有雄性的世界吗?
文献中对在快速变暖事件的气候情景下温度依赖性决定性别爬行动物的未来存在重大担忧[科顿和韦德金德,2009;西内尔沃等人,2010;Boyle 等, 2014]。这种担忧是合理的,事实上,最近在野生鬃狮蜥种群中发现性别逆转率上升,表明极端气候甚至可能扰乱看似稳定的基因型性别决定系统[ Holleley 等,2015]。因此,性别比例偏差有可能成为比最初预期更广泛爬行动物种群灭绝的驱动因素。
如果全球气温升高,爬行动物种群是否会变得性别偏倚愈发严重,并因种群变为单性而面临灭绝风险?答案取决于三个方面:1)爬行动物改变筑巢地点和时间的能力;2)耐热性适应的能力;3)环境条件变化的速度。随着温度上升,爬行动物种群中某一性别(通常是雄性)的出现频率降低,自然选择将强烈 favor 能够产生较稀有性别的个体 [ Boyle et al.,2014]。因此,如果种群中存在温度依赖型性别决定阈值的可遗传变异,该阈值的进化便是一种有效的响应方式,可降低风险存在极端的性别比例偏差。有大量证据表明,在产生混合性别比例的温度下,温度依赖型性别决定(TSD)爬行动物种群中存在广泛的遗传变异,这通过估计广义遗传力、狭义遗传力和/或基因型‐环境互作得以发现[Bull 等, 1982;Janzen,1992;Rhen 和 Lang,1998;Janes 和 Wayne,2006;McGaugh 和 Janzen,2011; McGaugh 等,2011;Rhen 等,2011]。支持这些遗传力研究的是器官培养实验,即在关键温度下独立培养的胚胎成对性腺在性别表型上保持一致,表明在孤立培养时对某一性别具有先天倾向[Mork 等, 2014]。
尽管存在其他涉及母体效应的解释,例如在器官培养前通过卵黄将性激素跨代传递,但这些实验表明,在关键温度下存在性别特异性的遗传倾向,此时温度对其影响并不明确。最后,在P. vitticeps中,有低频率的雌性个体(<4.2%)表现出高于平均水平的性别逆转热阈值,导致其ZZ后代在高温孵化条件下不会发生雌性化[Holleley et al.,2015]。因此,爬行动物物种无疑具有一定的适应温度依赖型性别决定阈值的能力。然而,热耐受性的进化需要在连续世代中实现成功繁殖。如果环境变化速度极为迅速,则仍可能存在极端性别比例偏差和局部/物种灭绝的重大风险。
结论
性别逆转现象发生在5个主要的爬行动物分支中,涉及XY和ZW系统,以及卵生和胎生物种。尽管我们对爬行动物中这一现象的认识仍处于初级阶段,但显然,环境性别反转是一种普遍存在的生物学过程,可能在爬行动物进化中发挥重要作用。事实上,性别逆转和基因‐环境相互作用极有可能导致现存爬行动物性别决定模式表现出的高度可塑性。本文讨论的案例无疑只是冰山一角,随着我们进一步解析爬行动物性染色体的基因组内容,并对已知性染色体的爬行动物开展更多的孵化研究,将会有更多性别逆转案例被揭示出来。我们认为,目前清晰的爬行动物性别逆转实例看似稀少,这不太可能反映该现象的真实发生率确实很低。几乎可以肯定,这种低检出率是由于爬行动物中性连锁标记和已确认的性染色体序列的缺乏所致。当前最值得关注的研究方向是阐明性别逆转如何与一些基本的进化过程相互作用,例如性染色体的产生与消失、物种形成事件,以及在气候变化背景下种群存续能力的变化。

















 671
671

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









