



生态数据显示,兰花多样性热点地区的蕾丽亚族在巴西大西洋森林可能处于濒危状态
引言
巴西拥有极为丰富的种子植物多样性,其中约50%的物种集中于大西洋森林,且为特有物种(Forzza 等 2012,BFG 2015)。这一生物多样性热点地区正面临森林砍伐和生境破碎化的严重威胁(Myers 等 2000,利马等,2015)。大多数森林碎片面积小于50公顷,而自然保护区仅保护了残余森林的一小部分( Ribeiro 等 2009)。近年来,巴西大西洋森林的森林砍伐率有所上升(SOS 大西洋森林基金会/国家空间研究院 2017)。许多巴西州的森林残存比例较低,例如里约热内卢州和米纳斯吉拉斯州分别约为19%和10%(SOS大西洋森林基金会/ 国家空间研究所 2017)。伊塔蒂亚亚国家公园(PARNA Itatiaia)是一个面积约30,000公顷的大型成熟森林碎片,位于这两个州境内。兰科是巴西第二大植物科,约有2,500个物种,其中60%分布于大西洋森林(BFG 2015)。伊塔蒂亚亚国家公园包含了约10%的巴西兰花物种(Barberena 2010),并被视为保护优先区域(Ribeiro 等 2009)。蕾丽亚族为新热带类群,主要特征为顶生叶、顶生(偶见侧生)花序以及紧密、蜡质、侧向扁平花粉块(Pridgeon 等 2005)。在巴西,该亚族包含24个属和约350个物种,多数具有很高的观赏价值,还包括一些变种和天然杂交种(Pabst 和 Dungs 1975;BFG 2015)。蕾丽亚族中的21个属以及超过60%的巴西物种分布于大西洋森林,其中约三分之二的物种为该植物地理区的特有物种(《2020年巴西植物志》)。
根据植物区系调查和生态学研究得出的更新后的区域物种名录对于建立保护工作至关重要,在评估红色名录中的分类单元时也极为有用。这在大西洋森林残余林区以及历史上长期受到采集压力的类群(如兰科)中尤为如此。
有关种群数量的数据经常缺失,阻碍了保护工作(梅尼尼·内托等,2013;伦齐 等,2015;利马等,2015)。附生植物在附生基质植物上的空间分布及其与附生基质植物专一性程度的分析,对于理解维管附生植物的生态与保护至关重要(卡多索等,2016;旺格勒等,2015)。
本研究提供了关于伊塔蒂亚国家公园(PARNA Itataia)蕾丽亚族物种的生态信息,并讨论了威胁这些物种的主要因素及官方保护政策,重点包括种群数量、垂直分布和兰科‐附生植物特异性。
材料与方法
伊塔蒂亚亚国家公园位于巴西东南部(22°14’–22°27’S,44°34’–44°46’ W),地处曼蒂凯拉山脉地区(ICMBio 2013,2017)。该公园具有显著的海拔梯度(600–2,791 m),包含山地(500–1,500 m)和高海拔山地(高于1,500 m)湿润森林(维洛索等,1991;ICMBio 2017)。后者还分布有高海拔草原,为开阔潮湿的草本植被,生长在海拔1,700 m以上地区(萨福德,1999)(图1、 2)。伊塔蒂亚亚国家公园包含三种气候类型:Csb、Cwb和Cwa(根据柯本, 1948)(ICMBio 2012)。年平均气温为11.4°C,年降水量为2,400 mm,平均相对湿度为75.2%(布拉德,1956;IBDF 1982)。公园地图使用ARC‐GIS软件,版本10.2绘制。
野外工作于2008年3月至2009年9月进行,主要集中在历史上采集到大多数蕾丽亚族标本的山地群落。同时也考察了公园的缓冲区。标本被
根据古德斯‐布鲁尼等人(2002)的方法进行地理定位、干燥和压制成标本,并保存在RB标本馆和SP标本馆中。具体的采集地点被有意省略。
检查了ALCB、GUA、HB、R、RB、RFA、RBR、SP和SPF标本馆的材料 (缩写依据蒂尔斯(2017)),包括P.C. 波特和A.C. 布雷德在伊塔蒂亚亚地区采集的标本,因为这些博物学家曾在现今属于该公园多个区域进行采集。材料的鉴定依据帕布斯特和邓格斯(1975, 1977)、斯普伦格尔等人(1996)以及专家鉴定过的标本进行比对完成。
采用Pridgeon等人(2005)提出并由范登贝格(2014)修订的蕾丽亚族分类系统。地理分布和植物地理区数据基于BFG(2015)、戈瓦尔茨等人(2017) 和热带植物数据库(2017)。基物数据来源于野外工作以及对公园内采集标本标签的分析,有时也参考BFG(2015)。受威胁物种的信息来自梅尼尼·内托等人 (2013)和巴西植物志(2020)。我们采用韦希特(1992)的分类方法对附主进行垂直分层(层次),以2米为间隔,并确定以下分布类型:(a)专性分布, 当物种局限于一个层次时;(b)偏好性分布,当局限于两个层次时;(c)广布型,当出现在三个或更多层次时。
还检查了公园内发现的兰亚族物种及其附生基质植物的特异性。提供了兰科物种及其附生基质植物发生层的所有可用信息。树木标本由里约热内卢植物园研究所的专家鉴定,并通过与RB标本馆的标本进行比对确认。
自十九世纪末以来,伊塔蒂亚亚国家公园(PARNA Itatiaia)便开始系统地采集兰科标本。一些蕾丽亚族物种曾被认为在当地存在
如果在过去50年中未采集到标本,并且没有可用信息表明它们仍存在于公园中,则视为灭绝。最近在公园内寻找它们的尝试均未成功。
结果
在伊塔蒂亚亚国家公园,蕾丽亚族由分布在六个属中的33个物种代表(表1; 图3)。下文总结了该公园内蕾丽亚族物种的地理分布和保护信息。
- 二色卡特兰林德利(1836a,图版1919)
附生植物(BFG 2015)。巴西特有物种,分布于大西洋森林和塞拉多(BFG 2015)。该物种被评估为近危(梅尼尼·内托等,2013)。在伊塔蒂亚亚国家公园的山地森林中有记录,约100年前曾采集到两次标本。现推测已局部灭绝。
- 猩红卡特兰林德利(1836a,图版1919)
附生植物。特有物种分布于巴西大西洋森林,涵盖东南部和南部地区的所有州 (BFG 2015)。该物种被评估为无危(梅尼尼·内托等,2013)。在伊塔蒂亚亚国家公园,见于高海拔山地森林及高海拔草原内的森林飞地,海拔范围为1,700–2,400 米,但更常见于2,000–2,200 米之间。在公园多个区域均有发现,种群数量估计超过200株。其生长于附生基质植物的六个基底层(高度分别为0–2 米、 2–4 米、4–6 米、6–8 米以及10–12 米),观察到其生长在 Weinmanniaorganensis Gardner(Cunoniaceae科)、Calyp thranthes concinna DC.(桃金娘科)、Myrceugenia cf. ovata (Hook. & Arn.) O.Berg(桃金娘科)、 Myrceugenia cf. camp estris(DC.) D.Legrand & Kausel(桃金娘科)以及一些豆科物种上。
- 卡特兰·克里丝帕·林德利(1828年,图1172)
附生植物(BFG 2015)。巴西大西洋森林特有物种,分布于米纳斯吉拉斯州、 里约热内卢州和圣保罗州(BFG 2015)。该物种被评估为易危(梅尼尼·内托等, 2013)。在伊塔蒂亚亚国家公园,近100年前曾在一未知地点采集到一次标本, 现推测已局部灭绝。
- 卡特兰·佩林尼·林德利 (1838a, 图版2)
附生植物(BFG 2015)。巴西大西洋森林特有,分布于米纳斯吉拉斯州、圣埃 斯皮里图州和里约热内卢州(BFG 2015)。该物种被评估为易危(梅尼尼·内托 等,2013)。在公园内的山地森林(海拔600米)曾有记录,100多年前曾两次采 集到该物种。目前被认为局部灭绝。
- 卡特兰紫红变种(林德利和帕克斯顿)范登伯格(2008年,第10页)
附生植物(BFG 2015)。巴西大西洋森林特有,分布于东南部和南部地区的多 个州(BFG 2015)。被评估为易危(梅尼尼·内托等,2013)。在伊塔蒂亚亚国 家公园,近100年前曾在一未知地点采集到一次。现被认为局部灭绝。
- 卡特兰·斯科菲尔德安娜·赖兴巴赫子 (1882年, 第808页)
附生植物(BFG 2015)。巴西大西洋森林特有,分布于圣埃斯皮里图州和里约 热内卢州(BFG 2015)。该物种属于极危等级(梅尼尼·内托等,2013)。在伊 塔蒂亚亚国家公园的山地森林中有记录,但仅在100多年前采集到一次。推测已 局部灭绝。
- 霍氏密花兰 钩(1830年,图版3013)
附生植物,偶为岩生。巴西特有物种,分布于伯南布哥州至南里奥格兰德州的大 西洋森林和塞拉多植物地理区(BFG 2015)。该物种的保护状况尚未评估。在 研究区域中,安蒂斯卷瓣兰(Encyclia atens)可能是与E idendrum secundum Jacq. 和Sca hyglottis modesta (Rchb.f.) Schltr. 同样最常见的蕾 丽亚族物种之一。该物种局限于山地森林,但见于多个地点,海拔范围为750– 1,150米。安蒂斯卷瓣兰生长于多种附生基质植物上,包括漆树科、番荔枝科和柏 科的物种,楝科的卡布腊莱亚·坎赫拉纳 [Vell.] Mart.),以及桃金娘科的Cam omanesia guaviroba[DC.] Kiaersk. 和细脉蒲桃 基尔斯基(Myrcia tenuivenosa Kiaersk.),其分布涵盖六个层次(高0–2米、2–4米、4–6米、6–8米、8–10米和10–12米)。
- 杏黄贝母兰 (1836年b, 图版1867)
附生植物。分布于南美洲,从厄瓜多尔、秘鲁和玻利维亚至巴西大西洋森林(从 东北部至南部地区)(BFG 2015;Govaerts 等人 2017)。该物种在巴西的保护 状况尚未评估。在伊塔泰亚国家公园(PARNA Itataia),E.armeniacum 生 长于山地群落中河流或小径边缘,海拔800–1,225米。该物种相对常见,已在五 个层次(0–2米、2–4米、4–6米、6–8米和8–10米高)以及多个被子植物科的多 种附生基质植物上发现。
- 小鸟贝母兰 林德利 (1841年a, 第85页)
岩生。广泛分布于南美洲,包括厄瓜多尔、秘鲁、玻利维亚以及巴西几乎所有地 区(北部地区除外),在这些地方均有发现
大西洋森林和塞拉多植物地理区(BFG 2015;Govaerts 等人 2017;Tropicos 2017)。在巴西,其保护状况尚未被评估。在研究区域,该物种仅发现于山地森 林中,但最近一次采集距今已超过50年,且仅记录于海拔800至1,000米的坎波贝 洛河沿岸。推测其已局部灭绝。
- 伊波芦荟·坎波斯‐波尔托伊·巴伯雷纳 巴伯雷纳 & 冈萨加 (2016, p. 227)
附生植物。特有物种,仅分布于里约热内卢州的伊塔蒂亚亚国家公园( Barberena 和 Gonzaga 2016)。Barberena 和 Gonzaga(2016)未对这一新 发现的物种进行评估,但指出火灾和非法采伐植物是其保护方面主要的本地威胁。该物种仅分布于海拔1,840米的高海拔山地森林。据Barberena和Gonzaga( 2016)记载,在研究区域内仅发现三个个体,它们彼此靠近(< 5 m),位于两 株附生基质植物(0–2米高)的基部层次。在评估该物种的保护状况之前,必须 对伊塔蒂亚亚国家公园内的种群数量进行充分调查。
- 伊波芦荟·克洛丽努姆·巴博萨·罗德里格斯 (1881, p. 139)
附生植物。特有于巴西大西洋森林和塞拉多植物地理区,分布于巴伊亚州及东南 地区(BFG 2015)。该物种被列为无危(梅尼尼·内托等,2013)。在研究区域, 该物种近35年来未被采集到标本。然而,最近在不同于最早采集记录的不同地点 发现了两个个体。该物种生长于海拔1,000–2,350米的山地森林和高海拔山地森 林中阴暗潮湿的生境,仅分布于附生基质植物最基底层(0–2米高处)。
- 库氏贝母兰 贝特曼 在 胡克(1867,图版5654)
附生植物。巴西大西洋森林特有物种,分布于东南地区的所有州。该物种的保护 状况尚未评估。该物种在研究区域仅局限于山地森林,且其生态特征尚无相关信 息。来自伊塔蒂亚亚国家公园的最新标本距今已有十余年,但采集记录十分稀少。
- 线茎贝母兰 林德利 (1853, 第101页)
附生植物。巴西大西洋森林特有物种,分布于巴伊亚州及东南地区的所有州。其 保护状况尚未评估。该物种局限于研究区域海拔800–950米的山地植被中,生长 在基底层(高2–4米)的被子植物附主上。该物种在该地区已超过95年未被采集 到,但在本研究期间发现了两个个体。目前令人担忧的是坎波贝洛河沿岸旅游业 的快速发展。
- 亨申氏贝母兰 巴博萨·罗德里格斯 (1881, 第147页)
附生植物。为巴西大西洋森林特有物种,分布于东南部和南部地区的五个州(米 纳斯吉拉斯州、里约热内卢州、圣保罗州、巴拉那州和圣卡塔琳娜州)(BFG 2015)。该物种被评估为濒危(梅尼尼·内托等,2013)。在伊塔蒂亚亚国家公 园较为常见,当地种群数量估计超过100个个体。其分布于坎波贝洛河流域,海 拔750–850米,生长在树参属 某种(五加科)和细脉蒲桃 基尔斯基(桃金娘科) 的基部和中间层(高2–4米、4–6米和6–8米)。
- 宽唇贝母兰 林德利 (1841b, p. 77)
附生植物。在巴西呈间断分布,见于亚马逊地区(亚马孙州和帕拉州)以及大西 洋森林的伯南布哥州和巴伊亚州,和东南部及南部地区的所有州(BFG 2015)。尽管波尔图 (1915) 记录了该物种在海拔约1,300米区域的存在,但仅发现一份90 年前采集的标本。本研究期间,在公园的缓冲区发现了附生个体,表明该物种可 能仍存在于该保护单元内(主要在邻近地区)。
- 马蒂凯拉兰姆贝母(1940,图版5)
附生植物。巴西大西洋森林特有物种,分布于东南地区的所有州(BFG 2015)。该物种的保护状况尚未评估。在公园内仅见于高海拔山地森林,生长在窄叶南洋 杉(Bertol.)孔策上,海拔范围为1,800–2,200米。
- 黄绿贝母兰巴巴罗德里格斯(1881,第140页)
附生植物。巴西大西洋森林特有物种,分布范围从伯南布哥州至南里奥格兰德州, 但塞尔希培州除外(BFG 2015)。该物种被评估为无危(Menini Neto 等, 2013)。在伊塔蒂亚亚国家公园的山地群落(海拔750米)中受限分布,生长于 坎波贝洛河岸边,位于被子植物附主基底层(高2–4米)处。最近仅发现一个个 体。在研究区域可能受威胁。
- 巴拉那贝母兰巴巴罗德里格斯(1881,第139页)
附生植物,罕见的岩生植物。特有于巴西塞拉多和大西洋森林,分布范围从巴伊 亚州至圣卡塔琳娜州(BFG 2015)。其保护状况被评估为无危(Menini
内托等人 2013)。该物种出现在公园的多个地点,但仅存在于山地植被中(海拔 750–1,250米),且始终靠近水道。它生长在番荔枝科和桃金娘科物种的基部和 中间层(高0–2米、2–4米和6–8米),并观察到几乎所有的花都发育成果实。
- 繁衍附生兰 巴博萨·罗德里格斯 (1877, 第61页)
附生植物。巴西大西洋森林特有,分布于伯南布哥州至南里奥格兰德州(BFG 2015)。被评估为无危(梅尼尼·内托等,2013)。在伊塔蒂亚亚国家公园的两 个地点有分布,海拔800至900米,生长于细脉蒲桃 基尔斯基(桃金娘科)的基 部和中间层(高0–2米、2–4米和4–6米)。然而,仅发现了六个个体。在研究区 域可能受威胁。
- 拟异形附生兰 赫内和施莱希特(施莱希特,1925年,第71页)
附生植物。巴西大西洋森林特有,分布于东南部和南部地区(圣埃斯皮里图州除 外)(BFG 2015)。其保护状况尚未评估。在伊塔蒂亚亚国家公园内,该物种 局限于海拔815–915米的山地群落,分布在多个游客到访的地点以及公园道路沿 线。它生长在梭罗树属物种(桑科)及其他被子植物附主上,位于三个层次(高 0–2米、2–4米和4–6米)。
- 多枝附生兰 雅克林 (1760年, 第29页)
附生植物,罕见岩生。广泛分布于美洲,从墨西哥到巴西南部。在巴西,E. ramosum 出现在亚马逊、塞拉多和大西洋森林植物地理区(BFG 2015; Govaerts 等人 2017)。其保护状况已
未被评估。在公园内,E. ramosum 出现在多个地点的海拔 750–1,100 米处,生 长于附生基质植物的基部和中间层(高度为 0–2 米、2–4 米、4–6 米和 6–8 米), 尤其见于番荔枝科和柏科物种以及杯盖花属(无患子科)上。
- 硬茎贝母兰 雅克林 (1760年, 第29页)
附生植物或岩生植物(BFG 2015)。广泛分布于美洲,从美国到阿根廷( Govaerts 等人 2017)。在巴西,该物种遍布全国(除皮奥伊州和北里奥格兰德 州外),分布于亚马逊、卡廷加、塞拉多和大西洋森林植物地理区(BFG 2015)。该物种的保护状况尚未被评估。在伊塔蒂亚亚国家公园,E. rigidum 记录于山 地森林中,曾在海拔900米的某未知地点采集到一次。此外,该物种已超过100年 未再被采集,推测在研究区域内已灭绝。
- 石上贝母兰 林德利 在 胡克 (1841b, 第84页)
附生植物(BFG 2015)。原产于南美洲,从圭亚那至巴拉圭(Govaerts 等人 2017)。在巴西,该物种呈间断分布,见于罗赖马州的亚马逊地区以及从塞阿拉 州到圣保罗州的塞拉多和大西洋森林植物地理区(BFG 2015)。其被评估为无 危(梅尼尼·内托等,2013)。在伊塔蒂亚亚国家公园,该物种曾于山地植被中 采集到,但仅记录一次且距今已超过75年。被认为局部灭绝。
- 单序贝母兰 雅克林 (1760年, 第29页)
陆生或岩生,偶为附生植物。分布遍及美洲,从美国至巴西南部(Tropicos 2017)。在巴西亦有广泛分布,见于亚马逊、卡廷加、塞拉多和大西洋森林植物 地理区(BFG 2015)。该物种被列为无危(梅尼尼·内托等,2013)。
Epidendrum secundum 在整个20th世纪均有采集记录,在研究区域中因其数 量丰富且分布广泛而引人注目。其生长海拔为750–2000米,主要位于河岸或路 边。该物种生长于基底层(高0–2米和2–4米)的棕榈科及其他被子植物附主上。
Epidendrum secundum 是伊塔蒂亚亚国家公园中唯一一种也能在土壤中生长 的蕾丽亚族物种。
- 球穗羊耳蒜(Reichenbach f. in Vriese 等人,1859年,第333页)
附生植物(BFG 2015)。分布于美国至热带美洲(Govaerts 等人 2017)。在巴 西,Epidendrum strobiliferum 分布广泛,见于亚马逊、塞拉多和大西洋森林 植物地理区,从北部(罗赖马州)到南部(南里奥格兰德州)均有分布(BFG 2015)。该物种的保护状况尚未评估。Epidendrum strobiliferum 曾记录于伊 塔蒂亚亚国家公园的山地森林中,波尔图 (1915) 提及其出现在约海拔900米处。然而,仅发现一份标本,且采集于70年前。该物种被认为已局部灭绝。
- 三指羊耳蒜(Lindley,1838b,第46页)
附生植物。南美洲,分布于厄瓜多尔、玻利维亚、秘鲁和巴西(Govaerts 等人 2017)。在巴西,仅分布于大西洋森林植物地理区,从帕拉伊巴至南里奥格兰德 州,被评估为无危(Menini Neto 等人
2013年)。在该公园内,三趾附生兰仅局限于海拔800–1050米的山地群落中的 少数地点。最近,在公园道路沿线发现了4株个体,生长于多种被子植物附主的 中层(高4–6米和6–8米)。值得注意的是,该物种已超过45年未被采集到标本。Barberena 91 (RB) 标本于2009年5月在人工栽培条件下开花,证实了该物种在 该区域的存在。该物种在研究区域可能受威胁。
- 林德氏贝母兰 (1838b,第50页)
附生植物(BFG 2015)。特有于巴西塞拉多和大西洋森林植物地理区,分布于 联邦区、阿拉戈斯州、巴伊亚州以及东南部和南部地区(BFG 2015)。该物种 被评估为无危(梅尼尼·内托等,2013)。在伊泰亚国家公园,囊状附生兰记录 于海拔1000米的山地森林,95年前曾在此采集到标本。现推测已局部灭绝。
- 伊莎白兰 (1877年,第76页)
附生植物或岩生(BFG 2015)。特有于大西洋森林,分布于巴西的米纳斯吉拉 斯州、里约热内卢州、圣保罗州和巴拉那州,以及巴拉圭和阿根廷(BFG 2015; Govaerts 等人 2017)。该物种被评估为易危(梅尼尼·内托等,2013)。在公园 内,伊诺佩鲁姆·维吉纳利斯记录于山地群落(海拔1300米),70年前曾采集到 一次(一个无性繁殖标本)。目前被认为局部灭绝。
- 阿勒曼类合萼兰(赫内)W.E.希金斯 (1997[1998], p. 376)
附生植物。特有物种,分布于巴西东南地区的巴西大西洋森林(BFG 2015)。该物种的保护状况尚未评估。拟宿柱兰在公园多个地点被发现,海拔800–1,100 米,生长于柏科物种、白桐树属(大戟科)、杯盖花属(无患子科)、某些豆科 物种以及其他被子植物附主的中上层(6–8米、8–10米和10–12米高)处。
- 筒距拟宿柱兰 (Lindley) W.E.Higgins (1997[1998], p. 377)
附生植物(BFG 2015)。特有物种,分布于托坎廷斯州(北部地区)、巴伊亚州 (东北地区)以及东南地区的所有州的大西洋森林和巴西塞拉多植物地理区( BFG 2015)。该物种的保护状况尚未评估。筒距拟宿柱兰在研究区域仅被采集 过一次,距今已超过100年,但采集地点未知。该物种被认为局部灭绝。
- 芳香拟宿柱兰 (Swartz) W.E.Higgins (1997[1998], p. 377)
附生植物(BFG 2015)。分布于美洲,从墨西哥至巴西南部(Govaerts 等人 2017)。广泛分布于巴西亚马逊地区和大西洋森林植物地理区(BFG 2015), 其保护状况尚未评估。尽管波尔图 (1915) 提到在伊塔蒂亚亚国家公园海拔850至 1,200米处采集到标本,但在该地区仅发现一份采于100多年前的标本。该物种被 认为局部灭绝。
- 厚萼合萼兰(克洛茨施)基隆和V.P.卡斯特罗 (2003, 第174页)
附生植物。特有物种,分布于巴伊亚州、米纳斯吉拉斯州、里约热内卢州和 圣保罗州的巴西塞拉多和大西洋森林植物地理区(BFG 2015)。该物种的保护状 况尚未评估。厚萼拟宿柱兰在伊塔蒂亚亚国家公园多个地点被采集,海拔800– 1,200米。其生长于文氏卡利普特兰瑟斯(Spreng.)D.Legrand(桃金娘科)及 其他被子植物物种上,位于基部和中间层次(高0–2米、2–4米和4–6米)。
- 纤巧桨唇兰(小赖兴巴赫)施莱希特(1926,第46页)
附生植物或岩生植物。广泛分布于中美洲、加勒比地区和南美洲(Govaerts 等 人 2017)。在巴西,该物种分布于亚马逊、卡廷加和大西洋森林植物地理区,从 北部地区的亚马孙州到南部地区的南里奥格兰德州(BFG 2015)。该物种在巴 西的保护状况尚未评估。纤巧桨唇兰仅见于公园内的山地森林,在多个地点均有 发现,通常位于潮湿生境,尤其是河流附近,海拔范围为750–1,200米。种群数 量估计超过200个个体。其生长宿主包括桤叶树属(桤叶树科)、瓜维罗巴坎波 马内西亚(DC.)基尔斯基(桃金娘科)、中间阿梅乌瓦马蒂乌斯(茜草科)和 杯盖花属(无患子科),以及一些番荔枝科和豆科物种。植株多见于中上层(高 4–10米),但也见于2–4米高度处。
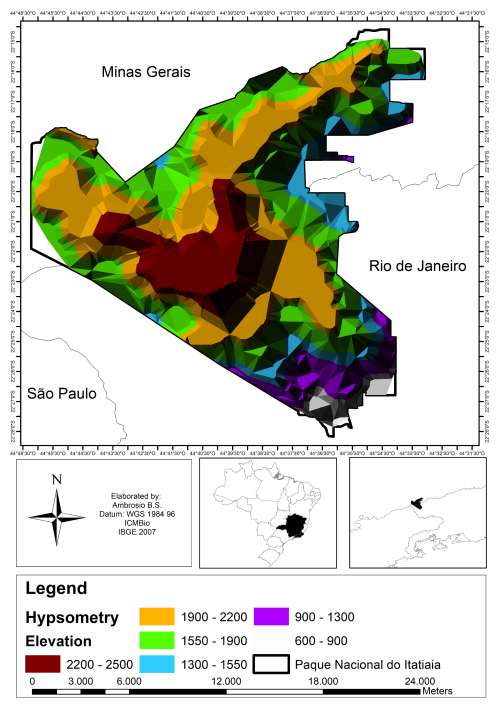


表1. 巴西东南部伊塔蒂亚亚国家公园的蕾丽亚族物种。
| 物种 | 凭证标本 |
|---|---|
| 双色卡特兰 | P.C. 波特 67 (RB) |
| 朱红卡特兰 | F.F.V.A. 巴伯雷纳 127 (RB) |
| 卷瓣卡特兰 | P.C. 波特 s.n. (RB 206766) |
| 佩林卡特兰 | P.C. 波尔图 无编号 (RB 8193) |
| 卡特兰紫红变种 | P.C. 波尔图 无编号 (RB 2067668) |
| 斯科菲尔德卡特兰 | P.C. 波尔图 68 (RB) |
| 宽萼羽唇兰 | F.F.V.A. 巴伯雷纳 35 (RB) |
| Epidendrum armeniacum | F.F.V.A. 巴伯雷纳 89 (RB) |
| 小鸟树兰 | P. C. 波尔图 1041 (RB) |
| ## 讨论 |
巴西的蕾丽亚族多样性最丰富的地区是大西洋森林(《2020年巴西植物志》),该植物地理区的新物种已被频繁描述(参见Barberena和Gonzaga,2016)。伊塔蒂亚亚国家公园的蕾丽亚族物种丰富度高于其他已开展该亚族分类学研究的巴西大西洋森林残余区域(例如Barbero,2007;Forster和Souza,2007;Romanini和Barros,2007;Nardy等,2016)。
伊塔蒂亚亚国家公园中大部分蕾丽亚族物种(73%)仅分布于山地植被中,而有四个物种(12%)局限于高海拔山地森林,两个物种(绿花树兰和 Epidendrum secundum )则出现在这两种植物群落中。对于研究区域,卷瓣卡特兰、紫纹兰和筒距拟宿柱兰这三个物种缺乏海拔数据。该公园的低海拔地区因19世纪和20世纪早期建立的咖啡种植园、畜牧业及殖民地聚居地而遭受严重破坏(托姆津斯基等人,2012年)。因此,海拔700至1200米的山坡发生了大规模森林砍伐(巴西生物多样性保护研究所 2012)。近几十年来,火灾、历史土地争端、非法采伐植物以及外来物种引入已成为植物保护的主要威胁(IBDF 1982)。布拉德(1956年)指出,火灾减少了伊塔蒂亚亚国家公园的树木种群,并促进了草本和灌木的生长。这种变化对兰花尤其是蕾丽亚族物种显然是不利的,因为它们通常需要高湿度环境和遮荫。关于上部山地植被和高海拔草原,火灾是主要威胁(莫里姆 2006;托姆津斯基等人,2012年)。2008年7月观察到这两类植被均发生火灾(F.F.V. 巴伯雷纳,个人通讯),但由于缺乏有关分布地点和种群规模的历史数据,尚无法评估其对兰花造成的影响。就“附生现象 ×层次”关系而言,没有任何物种局限于单一层次。 Epidendrum secundum 可归类为偏好性物种,另有11个物种被视为广布种。在伊塔蒂亚亚国家公园,大多数 E. secundum 标本为陆生,但偶尔,该物种会作为附生植物生长在湿度较高的附生基质植物的两个基部层次上。由于观察到的个体数量较少(< 10),埃皮登德鲁姆·坎波斯‐波尔图、 E. chlorinum 、蕨茎附生兰、黄绿附生兰和三趾附生兰被记录为仅出现在单一层次中。然而,对于研究区域内的许多蕾丽亚族物种而言,由于个体数量较少以及适合的附生基质植物有限,无法评估“附生现象 ×附生基质植物”之间的关系,从而难以进行适当的特异性统计分析。兰花通常对寄主树木没有特异性,但某些兰花与特定科(如棕榈科或山榄科物种)具有显著的特异性关联(特伦布莱等人 1998;梅代罗斯和贾尔丁 2011;巴贝雷纳等人 待出版)。尽管桃金娘科在大西洋森林中具有显著的物种丰富度(斯特曼等人 2009;BFG 2015),桃金娘科物种似乎是蕾丽亚族物种的良好附生基质植物,因其具备理想的特征(例如粗糙的树皮)。有七种兰科物种(卡特兰·科辛尼亚、 E. patens 、埃皮登德鲁姆·亨申尼、埃皮登德鲁姆·巴拉那种、繁衍附生兰、厚萼普洛斯特切亚、 S. modesta )被观察到生长于卡利普特兰塞斯属、坎波马内西亚属、蒲桃属和美洲蒲桃属的物种上。
PARNA伊塔蒂亚亚的六种蕾丽亚族物种此前在巴西被评估为受威胁物种:卡特兰·卷瓣种、卡特兰·佩林种、紫纹兰和伊诺佩鲁姆·维吉纳利斯被列为易危;埃皮登德鲁姆·亨申尼为濒危;卡特兰·肖菲尔德种为极危。此外, C. bicolor 被评定为近危。超过80%的卡特兰属物种为巴西特有,约三分之一的物种为大西洋森林特有物种(BFG 2015)。该属具有重要的园艺价值,是最广泛栽培和杂交的兰花类群之一(克鲁斯等,2003;Pridgeon 等 2005)。然而,卡特兰属物种通常在森林中密度较低,过度采集压力已导致其数量减少和多个种群的灭绝(克鲁斯等,2003;Pridgeon 等 2005)。在巴西,有41种卡特兰属物种受到威胁(梅尼尼·内托等,2013)。
卡特兰属无疑是伊塔蒂亚亚国家公园中最受威胁的属。在该区域的物种中,仅卡特兰·科辛尼亚近年来被采集到,并形成了较大的种群。其余五个物种在95至105年前仅被采集过一到两次。显然,它们大多数曾出现在海拔600米处一个名为本菲卡的地点,该地位于公园主入口附近,在20世纪初期经历了多次人为干扰。由于这些物种在伊塔蒂亚亚国家公园内具有狭域分布特性,再加上其突出的观赏价值,长期以来该地区的卡特兰植株很可能被无节制且秘密地采挖。尽管这些物种可能已经局部灭绝,但我们仍认为在一些难以到达或尚未探明的狭小区域中可能存在小型种群。同样,小鸟树兰、 E. rigidum 、 E. saxatile 、球果状附生兰、囊状附生兰、伊莎贝拉·维尔吉纳利斯、筒距拟宿柱兰和芳香普洛斯特切亚在过去50年中也未被采集到。这些物种主要记录于坎波贝洛河沿岸,而该河流夏季常发生洪水,且河岸地带存在生态旅游活动的影响。根据所采用的标准,有13个物种可被视为局部灭绝,约占伊塔蒂亚亚国家公园内蕾丽亚族物种总数的40%。此外,还有另外三个物种(黄绿附生兰、繁衍附生兰和三趾附生兰)在研究区域中也可能受到威胁。
另一方面,坎波斯‐波尔图附生兰最近被描述,为伊塔蒂亚亚国家公园特有物种。此外,本研究还发现了数十年来未曾采集到的物种(例如 E. chlorinum 、蕨茎附生兰、黄绿附生兰、繁衍附生兰、 E. ramosum 、三趾附生兰、阿勒曼状普洛斯特切亚和厚萼普洛斯特切亚)。然而,这些物种种群数量较小(通常为< 10个个体),因此亟需采取保护措施以确保其栖息地得到有效保护。亨申氏贝母兰和在本研究期间,首次在公园内采集到 E. pseudodifforme 。该种群出现在多个地点,且种群数量相对较大。在缓冲区的野外工作也具有重要意义,因为发现了 E. latilabre ,这表明它很可能也存在于公园内。
通常,形成较大种群的物种(如 E. secundum 和 S. modesta )在美洲具有较广的地理分布,或在巴西广泛分布(如 E. patens )。本研究结果与 Forster 和 Souza(2007)提供的数据一致。这些作者指出, Encylia patens 、 Epidendrum armeniacum (广泛分布于南美洲)和 E. secundum (遍布美洲)在卡帕拉奥地区有较好的代表性。相反,Forster 和 Souza(2007)将 C. bicolor 、 E. chlorinum 、 E. saxatile 以及至少其他六个分类单元归为稀有或易危物种,原因是发现的个体数量稀少,或近期未采集到标本,尽管已有多次尝试寻找它们。
Nardy 等(2016)强调人类活动影响是蕾丽亚族物种保护方面的一个令人担忧的问题。由于分布区面积缩减和基因库减少而导致的局部种群丧失,是对当前生物资源损失的一个重要警示信号(Ceballos 和 Ehrlich 2002)。鉴于大西洋森林不同残存林区均显示出蕾丽亚族物种局部受威胁比例较高的相似结果,并结合《巴西植物红色名录》中提供的数据(梅尼尼·内托等,2013),我们可以提出应对这一不利状况的策略。在此背景下,我们强调蕾丽亚族在巴西东南部大西洋森林残存林区中的高度多样性及其重要性,其中包括特有物种或已被列为受威胁的物种,以及许多物种的狭域分布特征。所有这些因素都支持保护大西洋森林庇护所的必要性。管理措施可与伊塔蒂亚亚国家公园负责人进行讨论,基于本研究中披露的科学信息实施,例如物种所处的偏好性生境、植物群落类型和海拔、生活型以及与区域分布数据相关的基质特性(主要是发生层)。
然而,首先需要回答一些问题。一种植物物种在什么情况下应被视为局部灭绝?在判断一种植物物种是否局部受威胁时,是否存在一个种群个体数量的最低阈值需要考虑?如何识别和衡量兰科物种的这种状况?在一个地区长达半个世纪甚至一个世纪未采集到个体,是否足以认定该物种在此地局部灭绝?当一个物种出现在保护单元的缓冲区时,是否应将其列为该法定保护区内的灭绝物种?
当涉及附生植物,尤其是兰花时,这些问题更加突出。巴罗斯(2007)指出,兰科植物在植物区系中尤其受到威胁,原因如下:(1)依赖寄主植物可获得性;(2)许多兰花对传粉者(特别是鸟类和昆虫)的依赖,直接影响其种群的繁殖和维持;(3)与某些真菌物种建立的相互关系,主要促进幼苗发育;(4)通常具有狭域分布;(5)爱好者和贸易商的采集压力,导致频繁的非法活动。伊塔蒂亚亚国家公园的蕾丽亚族物种可能面临所有这些因素,但在本次野外工作中仅观察到第(1)和第(4)项因素,而第(5)项因素被认为由于该类群物种的观赏价值,在20世纪期间已长期存在。除了明显的威胁(如火灾、森林砍伐)外,还需考虑其他影响兰花准确评估的因素:(6)种群结构研究表明,幼年阶段存在高死亡率,从而导致种群结构发生变化从聚集模式转变为随机空间分布模式(苏扎等人 2012);(7)附生兰花种群生长缓慢,再生率低;根据森哈斯 1993 的研究,兰花从萌发到首次开花的时间可从2½年到15年不等;(8)由于难以进入冠层这一层次,局限于树冠的植物经常未被采集或被错误鉴定。
在区域层面应用IUCN红色名录的类别和标准可能不适用于地缘政治边界,特别是对于地理范围过度受限的地区,因为无法确保该地区的种群与区域外的同种群隔离(IUCN 2012)。因此,在区域层面上,灭绝风险的估计往往不准确(IUCN 2012)。出于这一原因,还需解决一些额外问题:何时应正式向决策者提示某物种在某个州或保护单元中可能受威胁?应在何种阈值以下启动保护工作(资金、后勤等)?这些问题对于用于生物多样性保护资源相对较少的发展中国家尤为重要。大多数巴西保护区,包括伊塔蒂亚亚国家公园,都存在缺乏永久管控和监测的问题,并且在应对自然干扰和掠夺性人类活动方面面临困难。最后,值得思考的是,附生兰花的区域灭绝是否会对本地生态系统服务造成显著影响?但具体会如何影响?

















 692
692

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









