







RNA在细胞代谢中的角色远超其传统的基因信息传递功能。通过多种化学修饰,RNA不仅参与基因表达的调控,还在细胞的代谢网络和免疫反应中发挥关键作用。分享一篇总结的比较好的细胞代谢中的RNA修饰文章
8种RNA修饰
八种 RNA 修饰(m6A、m6Am、m1A、m5C、ac4C、m7G、Ψ和A-to-I编辑)的化学结构、分布和分子功能。
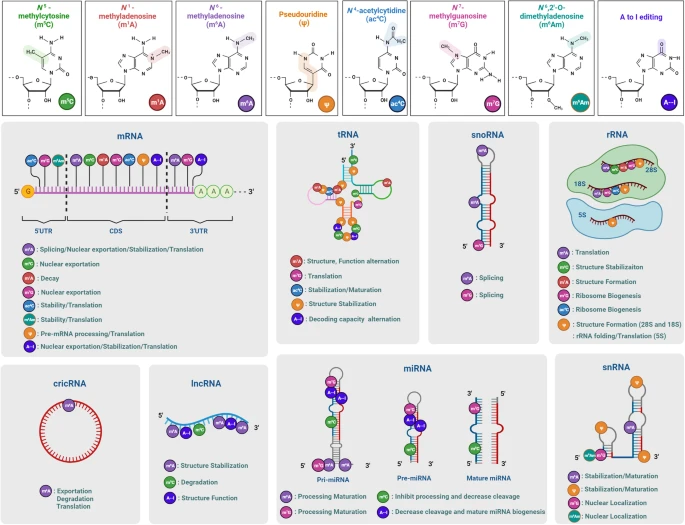
- m6A修饰:最广泛的RNA修饰之一,存在于mRNA、rRNA、snRNA、snoRNA、miRNA、lncRNA、circRNA和eRNA中。它由METTL3与METTL14组成的甲基转移酶复合物催化,影响RNA的翻译、稳定性、降解等过程。FTO和ALKBH5作为去甲基化酶,分别移除m6A和m6Am修饰。YTHDF家族蛋白及IGF2BP家族则作为“读者”蛋白,介导m6A修饰对RNA的稳定性和翻译等功能的调控。
- m6Am修饰:主要存在于mRNA的5’端和snRNA中。它由PCIF1催化,影响RNA的稳定性和翻译启动。METTL4也参与在U2 snRNA上催化m6Am修饰。FTO同样在此修饰上起到去甲基化作用。
- m1A修饰:主要发生在tRNA、rRNA、lncRNA和mRNA中,由TRMT61B、TRMT10C、TRMT6/61A等酶催化。它参与翻译的调节,并影响RNA的稳定性和功能。FTO、ALKBH1、ALKBH3和ALKBH7等去甲基化酶对m1A进行去甲基化处理,进一步调节其功能。
- m5C修饰:在tRNA和rRNA中,由NSUN家族酶催化。NSUN1、NSUN5等酶在28S rRNA上进行m5C修饰,而NSUN2则在多个tRNA上进行修饰。ALKBH1、TET1/2/3等去甲基化酶对m5C进行去甲基化,调节RNA的稳定性和翻译。
- ac4C修饰:存在于mRNA、tRNA和rRNA中,由NAT10等酶催化。它主要影响RNA的翻译效率,尤其是在tRNA和18S rRNA中的修饰,参与细胞应答和适应性反应。
- m7G修饰:广泛存在于mRNA、tRNA、rRNA和miRNA中,由METTL1等酶催化。它参与RNA的核出口、翻译起始和稳定性。RNMT和WBSCR22等酶对m7G修饰进行加帽作用,TGS1则在snRNA和snoRNA中将m7G进一步修饰为m2,2,7G。
- Ψ修饰:Ψ修饰,伪尿嘧啶,主要出现在rRNA中,由DKC1等酶催化。它通过形成伪尿嘧啶与其他RNA结构稳定性增强,从而在RNA功能精确性方面发挥作用,特别是在翻译过程中至关重要。
- A-to-I编辑:由ADAR家族的酶(如ADAR1、ADAR2)催化的,它通过将腺嘌呤(A)转变为肌苷(I),广泛影响mRNA的剪接、翻译效率和基因表达调控。ADAR3则不具有去氨基酶活性,但仍与编辑过程相关。
RNA 修饰的主要数据库
目前,RNA修饰数据库已建立15个,其中两个数据库集中于RNA修饰的生化特征,其余数据库旨在阐明其生物学作用,后者包括可逆RNA修饰数据库,可进一步分为综合性和类型特异性,以及不可逆RNA修饰数据库,即RNA编辑数据库
| 名称 | 描述 | URL |
|---|---|---|
| RNAMDB | 109种RNA修饰核苷酸的基本化学特征数据库 | https://mods.rna.albany.edu/ |
| MODOMICS | 最全面的RNA修饰途径资源 | http://modomics.genesilico.pl |
| m6A-Atlas | 一个用于揭示m6A表观转录组的综合知识库 | www.xjtlu.edu.cn/biologicalsciences/atlas |
| m7GHub v2.0 | 一个解读mRNA内部m7G表观转录组的定位、调控和发病机制的资源 | www.xjtlu.edu.cn/biologicalsciences/m7ghub |
| m5C-Atlas | 解码和注释m5C表观转录组的数据库 | https://www.xjtlu.edu.cn/biologicalsciences/m5c-atlas |
| MeT-DB v2.0 | 用于研究m6A的数据库,前一个版本是第一个关于m6A在转录组中的综合资源 | http://compgenomics.utsa.edu/MeTDB/ |
| RMBase v2.0 | 解读RNA修饰图谱的数据库,基于表观转录组测序数据 | http://rna.sysu.edu.cn/rmbase/ |
| REPIC | 一个m6A甲基组图谱,具有细胞系或组织特异性 | https://repicmod.uchicago.edu/repic |
| CVm6A | 一个用于探索和可视化全球m6A模式的数据库,涵盖不同细胞系 | http://gb.whu.edu.cn:8080/CVm6A |
| RMVar | 一个涉及RNA修饰的功能变异的数据库 | http://rmvar.renlab.org |
| RMDisease | 一个揭示与疾病相关的变异及其表观转录组扰动的数据库 | www.xjtlu.edu.cn/biologicalsciences/rmd |
| REDIdb | 一个专门的植物线粒体RNA编辑修饰数据库 | http://srv00.recas.ba.infn.it/redidb/index.html |
| RADAR | 一个严格注释的A-to-I RNA编辑数据库,涵盖人类、小鼠和果蝇 | http://RNAedit.com |
| DARNED | 一个集中在A-to-I编辑的人类RNA编辑数据库 | https://darned.ucc.ie/ |
| REDIportal | 最大且专门的A-to-I编辑数据库,涉及多种人类组织 | http://srv00.recas.ba.infn.it/atlas/ |
细胞代谢
细胞代谢是细胞维持生命活动的基础,而RNA修饰通过影响代谢途径的关键酶、转运蛋白以及转录因子的表达,调节着细胞的代谢过程。文章总结了四种代谢
- 葡萄糖代谢:细胞能量产生的核心,包括糖酵解、有氧氧化、糖原合成和糖异生等过程。在癌细胞中,糖酵解即使在有氧条件下也会优先于线粒体氧化磷酸化,这一现象称为Warburg效应。RNA修饰通过调节关键酶(如己糖激酶、乳酸脱氢酶)和葡萄糖转运蛋白的表达,参与糖代谢的调控。糖尿病和癌症等病理中,m6A修饰、FTO、METTL3等在调控葡萄糖代谢方面发挥重要作用。
- 脂质代谢:细胞膜的基本组成部分,也是能量储存的重要来源。在能量充足时,脂肪酸通过酯化储存在脂滴中,而在能量缺乏时,脂肪酸通过β-氧化生成ATP。脂质合成受固醇调节元件结合蛋白(SREBP1c)的调控。RNA修饰参与脂质代谢的多个环节,调节脂肪酸合成和分解等过程,影响细胞的能量平衡。
- 氨基酸代谢:氨基酸代谢涉及合成、降解以及调节氨基酸水平的关键途径。RNA修饰通过调节氨基酸代谢相关基因的表达,影响细胞的氨基酸合成、蛋白质合成及细胞内信号传导路径。
- 线粒体代谢:细胞能量的主要供应者,涉及氧化磷酸化和ATP的合成。RNA修饰通过影响与线粒体功能相关的基因的表达,如与ATP合成相关的酶,调节线粒体代谢和能量产生。
| 调控因子 | 疾病 | 目标 | 机制 | 代谢组别 |
|---|---|---|---|---|
| METTL3 | 肥胖 | FASN | 上调mRNA水平 | 脂质代谢 |
| CCND1 | 通过YTHDF2促进CCND1 mRNA降解 | 脂质代谢 | ||
| FTO | RUNX1T1 | 调控RNA结合能力 | 脂质代谢 | |
| CCNA2/CDK2 | 降低YTHDF2介导的mRNA衰变 | 脂质代谢 | ||
| YTHDF1 | NAFLD | MTCH2 | 促进翻译 | 脂质代谢 |
| METTL3/14 | NAFLD | ACLY/SCD1 | 上调基因表达 | 脂质代谢 |
| FTO | SREBP1/SCD1 | 通过m6A去甲基化上调基因表达 | 脂质代谢 | |
| ALKBH5 | LINC01468 | 稳定LINC01468,促进INPPL1降解 | 脂质代谢 | |
| YTHDF3 | PRDX3 | 促进翻译 | 脂质代谢 | |
| METTL14 | AS | p65 | 促进p65的表达 | 氨基酸代谢 |
| FTO | BLCA | PPARγ | 降低PPARγ表达并促进AMPK磷酸化 | 脂质代谢 |
| METTL14 | BRCA/HCC | CPT1B | 通过m6A水平上调CPT1B | 脂质代谢 |
| METTL5 | HCC | ACSL4 | 促进FAO | 脂质代谢 |
| FTO | EC | HSD17B11 | 增强翻译 | 脂质代谢 |
| YTHDF2 | CRC | DEGS2 | 促进降解,诱导脂质失调 | 脂质代谢 |
| IGF2BP2 | AML | MFSD2A | 稳定PRMT6 mRNA抑制MFSD2A表达 | 脂质代谢 |
| METTL3 | NSCLC | DCP2 | 加速降解 | 线粒体代谢 |
| METTL14 | CRC | miR-17-5p | 通过YTHDC2降解miR-17-5p | 线粒体代谢 |
| IGF2BP1 | GC | NDUFA4 | 稳定NDUFA4 mRNA增强氧化代谢 | 线粒体代谢 |
| FTO | ccRCC | PGC-1α | 上调PGC-1α表达 | 线粒体代谢 |
| METTL3 | BRCA | AK4 | 上调AK4,促进ROS生成 | 线粒体代谢 |
| FTO | ccRCC | SLC1A5 | 上调SLC1A5表达 | 氨基酸代谢 |
| YTHDF1 | CRC | GLS | 促进GLS的翻译 | 氨基酸代谢 |
| IGF2BP2 | AML | GPT2/SLC1A5/MYC | 稳定多个氨基酸代谢相关基因的mRNA | 氨基酸代谢 |
| METTL16 | AML | BCAT1/2 | 促进表达,调节支链氨基酸代谢 | 氨基酸代谢 |
免疫代谢
一般而言,免疫代谢涉及活化和静息免疫细胞之间的区别。前者的代谢方式类似于恶性细胞,Warburg效应,没有明显的OXPHOS,而后者从FAO和Krebs循环中获取能量。这里重点关注RNA修饰对不同免疫反应中免疫代谢的贡献。
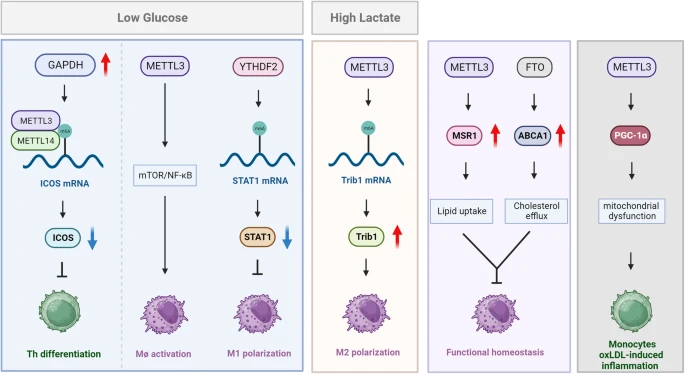
1. 抗肿瘤免疫
- 肿瘤免疫微环境(TIME):肿瘤细胞消耗大量葡萄糖和谷氨酰胺,形成不利的免疫细胞环境,导致免疫抑制。
- T细胞的葡萄糖代谢:T细胞在肿瘤微环境中受到糖酵解抑制,但VHL缺乏可诱导糖酵解酶GAPDH的表达,增强ICOS mRNA上的m6A修饰,抑制ICOS表达,从而影响Tfh细胞的分化。
- M1巨噬细胞的代谢适应:m6A修饰通过YTHDF2介导的STAT1 mRNA降解,抑制糖酵解相关基因的表达,进而影响M1巨噬细胞极化。
- 乳酸的免疫抑制作用:乳酸积累通过m6A修饰支持Treg细胞,促进免疫逃逸。
- 脂质代谢的影响:肿瘤细胞分泌的脂质对T细胞、树突状细胞和巨噬细胞产生有害影响,m6A甲基化通过调节脂质摄取平衡维持巨噬细胞功能。
2. 抗病毒免疫
- 病毒RNA的m6A修饰:m6A、m5C、ac4C、Ψ等RNA修饰调控病毒RNA的感知和信号传导,影响抗病毒免疫反应。
- 干扰素途径的调控:METTL3/14通过YTHDF2提高IFNB mRNA的周转率,加速病毒传播。
- ALKBH5与病毒传播:ALKBH5通过代谢重组促进病毒传播,影响病毒的复制。
- RNA修饰与免疫细胞功能:病毒感染通过改变RNA修饰影响免疫细胞功能,但与病毒感染的具体机制仍有待进一步研究。
3. 炎症和自身免疫性疾病
- m6A修饰在炎症中的作用:m6A修饰参与调节自身免疫性疾病和炎症反应。例如,METTL3在类风湿性关节炎(RA)患者中显著上调,与CRP和ESR等炎症标志物呈正相关。
- m5C在系统性红斑狼疮(SLE)中的作用:SLE中CD4+T细胞中m5C水平降低,NSUN2表达减少,影响炎症途径的基因表达。
- 树突状细胞依赖的炎症反应:m6A修饰通过调节糖酵解和树突状细胞迁移,影响免疫反应中的反馈控制。
- 巨噬细胞极化与代谢重编程:m6A甲基化通过增强HDGF的表达,调节M1巨噬细胞的糖酵解和脂质积累,进而影响动脉粥样硬化等疾病的发展。
- METTL3与PGC-1α在炎症中的作用:METTL3介导的PGC-1α mRNA的m6A修饰参与线粒体功能障碍和氧化低密度脂蛋白(oxLDL)诱导的单核细胞炎症。
RNA修饰与代谢和免疫疗法的联合应用正在成为治疗耐药性的重要策略。通过靶向m6A调节因子如METTL3、FTO等,研究发现可以克服肿瘤的化学耐药性并增强免疫治疗效果。例如,METTL3通过调节5-FU耐药性,FTO抑制剂能恢复对顺铂的敏感性。RNA修饰调节剂与现有抗癌药物的联合应用,如与免疫检查点阻断疗法(ICB)配合使用,已显示出改善肿瘤免疫反应的潜力。


















 454
454

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言











