




注:本文为 “趋同进化” 相关合辑。
英文引文,机翻未校。
中文引文,略作重排。
如有内容异常,请看原文。
Convergent Evolution
趋同进化
By: BD Editors
Reviewed by: BD Editors
Last Updated: April 28, 2017
Convergent Evolution Definition
趋同进化定义
Convergent evolution is the process in which organisms that are not closely related independently evolve similar features. Adaptations may take the form of similar body forms, colors, organs, and other traits that make up the organism’s phenotype.
趋同进化是非密切相关的生物体独立进化出相似特征的过程。适应可以采取构成生物体表型的相似身体形态、颜色、器官和其他特征的形式。
Convergent evolution creates analogous structures or “homoplasies”, those which have similar forms or functions between diverged species, but were not present in the common ancestor of the two. On the other hand, homologous structures, i.e., a specific organ or bone which appears throughout many different organisms, albeit often in a slightly different form or shape, can indicate a divergence from a common ancestor.
趋同进化创造了类似的结构或“同质体”,即那些在分化物种之间具有相似形式或功能,但不存在于两者的共同祖先中的结构。另一方面,同源结构,即出现在许多不同生物体中的特定器官或骨骼,尽管通常以略有不同的形式或形状出现,可以表明与共同祖先的分歧。
There are several circumstances that can result in convergent evolution. Often, convergence occurs when organisms are required to adapt to similar environmental conditions, such as in the evolution of thick water-retaining leaves and spines on cacti and Euphorbia species, which are adapted to tolerate conditions of extreme drought but are native to separate continents. It may also occur when two different organisms occupy a similar niche, for example, the cryptic green coloration of Emerald Tree Boas (Corallus caninus) from South America and Green Tree Pythons (Chondropython viridis) from Australia, both of which live high up in the canopy of similar rainforests and occupy a niche preying upon birds.
有几种情况会导致趋同进化。通常,当生物体需要适应类似的环境条件时,就会发生趋同,例如仙人掌和大戟物种上厚厚的保水叶子和刺的进化,这些物种适应了极端干旱的条件,但原产于不同的大陆。当两种不同的生物占据相似的生态位时,也可能发生这种情况,例如,来自南美洲的翡翠树蟒(Corallus caninus)和来自澳大利亚的绿树蟒(Chondropython viridis)的神秘绿色,它们都生活在类似雨林的树冠高处,占据了捕食鸟类的生态位。
Convergence of life cycle and behavioral traits, such as the similar social colony structures between Naked Mole-rats (Heterocephalus glaber) and many species of social bees and ants, can also take place in order to maximize the breeding success of individuals and within colonies. On a molecular level, the independent evolution of proteins and toxins has also occurred throughout many separate phyla; for example, sea anemones (Cnidaria), snakes (Vertebrates), scorpions (Arthropods), and cone snails (Molluscs) all produce neurotoxins which act similarly upon the neurotransmitter receptors of their prey.
生命周期和行为特征的趋同,例如裸鼹鼠(Heterocephalus glaber)与许多种类的社会性蜜蜂和蚂蚁之间的相似社会群体结构,也可以发生,以最大限度地提高个体和群体内的繁殖成功率。在分子水平上,蛋白质和毒素的独立进化也发生在许多不同的门中;例如,海葵(刺胞动物)、蛇(脊椎动物)、蝎子(节肢动物)和锥形蜗牛(软体动物)都会产生神经毒素,这些毒素对猎物的神经递质受体起作用相似。
Convergent evolution can also arise through mimicry complexes, in which organisms evolve to replicate the morphology of other species. This adaptation benefits the mimic either by way of protection when imitating the phenotype of an organism that is toxic or otherwise dangerous (Batesian mimicry), or allowing the mimic to exploit a resource or interaction by being mistaken for the model (Müllerian mimicry).
趋同进化也可以通过拟态复合体产生,其中生物体进化以复制其他物种的形态。这种适应有利于模仿者,要么在模仿有毒或其他危险的生物体的表型时提供保护(贝茨拟态),要么允许拟态者通过被误认为模型来利用资源或相互作用(苗勒拟态)。
The process of convergent evolution is in contrast to divergent evolution, whereby species which are closely related evolve different traits, and parallel evolution, in which similar traits develop in related, although distinct, species from a common ancestor but from different clades.
趋同进化的过程与发散进化形成鲜明对比,发散进化是密切相关的物种进化出不同的性状,而平行进化则在来自共同祖先但来自不同进化枝的相关但不同的物种中发展出相似的性状。
Examples of Convergent Evolution
趋同进化的例子
Convergent Evolution of Wings
翅膀的趋同进化
A widespread example of convergent evolution is the evolution of wings and powered flight in birds, bats, and (now extinct) pterosaurs, each of which belong to a different class of organism and therefore have very distant common ancestors.
趋同进化的一个广泛例子是鸟类、蝙蝠和(现已灭绝的)翼龙的翅膀和动力飞行的进化,它们中的每一种都属于不同类别的生物,因此具有非常遥远的共同祖先。
Fossil evidence has determined that flight evolved in pterosaurs (flying reptiles of the late Triassic period) around 225 mya and in birds around 150 mya, while mammalian bats evolved wings around 50-60 mya. The evolution of powered flight has only happened once in each of these lineages, although there are certain organisms, for example, ostrich birds, which have subsequently reverted back to being flightless while retaining their wing structures.
化石证据表明,翼龙(三叠纪晚期的飞行爬行动物)的飞行进化程度约为 225 mya,鸟类的飞行进化程度约为 150 mya,而哺乳动物蝙蝠的翅膀进化程度约为 50-60 mya。动力飞行的进化在这些谱系中的每一个中只发生过一次,尽管有些生物体,例如鸵鸟,随后又恢复了无法飞行的状态,同时保留了它们的翅膀结构。
The different wing structures of birds, bats, and pterosaurs are each supported by a modified five-fingered limb. Each limb consists of a humerus, a radius and ulna, a thumb and finger bones, and is a homologous structure, containing the same bones that make up the limbs of many animals including humans, whales, and crocodiles; however, the shape of each bone differs greatly between each form.
鸟类、蝙蝠和翼龙的不同翅膀结构都由改良的五指肢体支撑。每个肢体由肱骨、桡骨和尺骨、拇指骨和手指骨组成,是一个同源结构,包含构成许多动物四肢的相同骨骼,包括人类、鲸鱼和鳄鱼;然而,每种形式之间每块骨头的形状差异很大。
An elongated fourth finger shapes the pterosaur wing, with the other digits used as claws. In birds, an elongated radius and ulna, as well as finger bones fused together for strength, support the wing. Finally, the wings of a bat differ in that they are formed of membrane that is stretched over four elongated fingers. The reason each of these different bone formations results in the same eventual wing shape is due to the basic physics of flight: wings that were shaped much differently would not allow an animal to fly.
细长的无名指塑造了翼龙的翅膀,其他手指用作爪子。在鸟类中,细长的桡骨和尺骨,以及融合在一起以获得力量的指骨支撑着翅膀。最后,蝙蝠的翅膀的不同之处在于它们是由拉伸在四个细长手指上的膜组成的。这些不同的骨骼结构中的每一个都导致相同的最终翅膀形状是由于飞行的基本物理原理:形状大不相同的翅膀不允许动物飞行。
Although birds and pterosaurs share a very distant common ancestor, and birds also share a common ancestor with bats, none of these ancestors had wings or were able to fly. In each of these lineages, the wing is therefore an analogous structure because the bones have been arranged differently in order to independently achieve a functionally similar structure.
尽管鸟类和翼龙有一个非常遥远的共同祖先,鸟类也与蝙蝠有一个共同的祖先,但这些祖先都没有翅膀或能够飞行。因此,在这些谱系中的每一个中,翅膀都是一个类似的结构,因为骨骼的排列方式不同,以便独立实现功能相似的结构。

The image above shows the different internal bone structures of wings in: 1) Reptilian pterosaurs (Pterosauria). 2) Mammalian bats (Chiroptera). 3) Birds (Aves).
上图显示了翅膀的不同内部骨骼结构:1) 爬行动物翼龙(Pterosauria)。2)哺乳动物蝙蝠(Chiroptera)。3) 鸟类(Aves)。
Convergent Evolution between Placental Mammals and Marsupials
胎盘哺乳动物和有袋动物之间的趋同进化
Placental mammals, which have offspring that undergo gestation within the uterus and are born fairly advanced, and marsupials whose offspring are born very immature and continue to develop within a pouch on the mother’s body, diverged from a common ancestor around 100 million years ago.
胎盘哺乳动物的后代在子宫内进行妊娠并出生时相当晚期,而有袋动物的后代出生时非常不成熟,并在母亲身体上的育儿袋内继续发育,它们在大约 1 亿年前与共同祖先分化。
Separated by the split of continents, mammals evolved to occupy niches in Europe, Africa, and America, while marsupials occupied similar niches in Australia and the surrounding islands; this history has produced many examples of convergent evolution.
由于大陆的分裂,哺乳动物进化到占据欧洲、非洲和美洲的生态位,而有袋动物则在澳大利亚和周边岛屿占据了类似的生态位;这段历史产生了许多趋同进化的例子。
Animals from each group developed similar analogous structures depending on factors such as their habitat, feeding habits, and locomotion requirements.
每组动物根据栖息地、摄食习惯和运动要求等因素发育出相似的类似结构。
Burrowing animals evolved into the mole and the marsupial mole, which have similar body shapes, claws for digging, and lack efficient eyesight.
穴居动物进化成鼹鼠和有袋动物鼹鼠,它们具有相似的体型、用于挖掘的爪子和缺乏有效视力。
The Thylacine (now extinct) filled the same niche as the wolf: an apex predator with sharp teeth, powerful jaws, and speed for successful hunting.
袋狼(现已灭绝)与狼占据了相同的生态位:一种顶级掠食者,拥有锋利的牙齿、强大的下巴和成功狩猎的速度。
Placental Flying Squirrels and marsupial Sugar Gliders both evolved from the same flightless, common ancestor which split around 65 million years ago. These two animals are extremely similar in appearance and behavior; they are approximately the same size, they have large eyes for foraging in the dark, they are coated with soft fur, and have light underbellies.
胎盘鼯鼠和有袋蜜袋鼯都是从大约 6500 万年前分裂的同一个不会飞的共同祖先进化而来的。这两种动物在外观和行为上极其相似;它们的体型大致相同,它们有大眼睛,可以在黑暗中觅食,它们覆盖着柔软的皮毛,下腹部很浅。
Through convergent evolution, they have also both evolved structures that enable them to glide between the treetops where they live. The wing-like structures are made of skin, which is stretched between the forelimb and hind limb, and do not enable powered flight. Nonetheless, it is theorized that these may be analogous structures that appear as a precursor to flight.
通过趋同进化,它们也进化出结构,使它们能够在它们居住的树梢之间滑行。翅膀状结构由皮肤制成,皮肤在前肢和后肢之间伸展,无法实现动力飞行。尽管如此,理论上,这些可能是作为飞行前兆出现的类似结构。


The images show the skin stretched between the limbs of a Sugar Glider and a Flying Squirrel, evolved to allow gliding motion.
这些图像显示了蜜袋鼯和鼯鼠四肢之间伸展的皮肤,进化为允许滑翔运动。
Some Other Examples of Convergent Evolution
趋同进化的其他一些例子
- The evolution of complex eyes in vertebrates, cephalopods (squid and octopus), and arthropods (crustaceans, insects, and spiders).
脊椎动物、头足类动物(鱿鱼和章鱼)和节肢动物(甲壳类动物、昆虫和蜘蛛)复杂眼睛的进化。 - The streamlined body shape of dolphins, sharks, and (extinct) ichthyosaurs.
海豚、鲨鱼和(已灭绝的)鱼龙的流线型体型。 - The evolution of echolocation in whales and bats.
鲸鱼和蝙蝠回声定位的进化。 - The paired shell shape of bivalve mollusks and brachiopods.
双壳类软体动物和腕足类动物的成对壳形状。 - The silk-producing ability of spiders, silk worms, silk moths, and weaver ants.
蜘蛛、蚕、蚕蛾和织布蚁的产丝能力。 - The long structures (tongues and beaks) evolved for collecting nectar in hummingbirds, bees, moths, and butterflies.
长结构(舌头和喙)是为了收集蜂鸟、蜜蜂、飞蛾和蝴蝶的花蜜而进化而来的。 - The evolution of eyespots on the wings of butterflies and the tails of fish.
蝴蝶翅膀和鱼尾巴眼点的进化。 - Spines on the bodies of echidnas (monotremes), hedgehogs (mammals), and porcupines (rodents).
针鼹(单孔目动物)、刺猬(哺乳动物)和豪猪(啮齿动物)身上的刺。 - Filter feeding in many whales (such as humpback and baleen), sharks (such as whale sharks and basking sharks), and manta rays.
许多鲸鱼(如座头鲸和鲸须)、鲨鱼(如鲸鲨和姥鲨)和蝠鲼的滤食性。 - The evolution of the woody stem in seed plants, horsetails, and trees.
种子植物、马尾和树木中木本茎的进化。 - The reef-building abilities of many sea life organisms such as corals, sponges, cnidarians, and bacteria.
许多海洋生物的珊瑚礁建造能力,如珊瑚、海绵、刺胞动物和细菌。
Related Biology Terms
相关生物学术语
- Divergent Evolution – The evolution and accumulation of different traits between groups, which results in the formation of new species.
发散进化——群体之间不同性状的进化和积累,导致新物种的形成。 - Vestigial Structures – A structure or attribute, which is present within an organism but has lost its ancestral function.
退化结构 – 一种结构或属性,存在于生物体内但已失去其祖先功能。 - Analogous Structures – An organ or structure, which is visually similar or performs the same function in two different species, although, is not present in their common ancestor.
类似结构 – 一种器官或结构,在视觉上相似或在两个不同的物种中执行相同的功能,尽管它们的共同祖先中不存在。 - Homologous Structures – An organ or structure in the body, which is inherited from a common ancestor between species.
同源结构 – 体内的器官或结构,遗传自物种之间的共同祖先。
原本不一样,现在长得像:趋同进化是个啥?
博物 2022 年 07 月 26 日 11:15 北京
这一期我们就来翻翻巨口鲨的族谱,追溯下巨口鲨的进化史。

巨口鲨属于滤食性鲨鱼,经过这么多期锲而不舍的念叨,大家应该都记住了。但是,提到滤食性的海洋生物,有没有感觉有那么一丝熟悉?是的,蓝色海洋中最大的生物蓝鲸,还有最大的鱼鲸鲨,都是滤食性的。一个是鲸、两个是鲨,为什么不同的生物会有同样的取食方式?想知道这个问题的答案,还得循其根本,从进化层面上去探究。
非亲非故,但是长得像
仔细想想,不止是鲸和鲨,我们在大自然中经常能够看到不同的物种具有相似特征的现象。这来自于生物进化的一种方式:趋同进化。
趋同进化,指的是两种或几种亲缘关系较远、没有较近的共同祖先的物种,由于相近的环境和生活方式,演化出相似的形态或结构特征。简单来讲,就是两种 “非亲非故” 的生物 “越长越像”。趋同进化常常发生在如下情境中:一种是当不同的生物需要适应相同的环境时;另一种情况是在不同的地理位置存在相同或相似的生态位时,不同的物种很可能会占据该生态位。生态位(ecological niche)这个概念是由美国野外生物学家与动物学家乔瑟夫・格林尼尔(Joseph Grinnell)在 1917 年首次提出的,生态位的概念更具体地解释了生物是环境塑造的,抽象概念生态位就像是一个模具,无论流入这个模具的材质是什么,最终凝聚的形态都总是类似的。
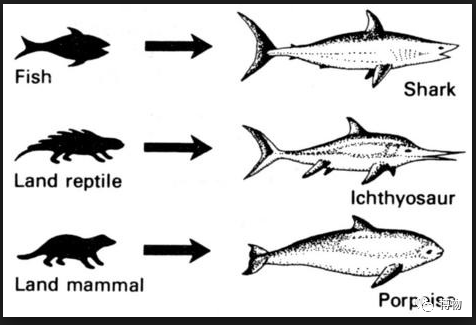
鲨鱼、鱼龙、鼠海豚的趋同进化
来源:https://ngtiankhean.com/
鲨鱼鲸鱼,入乡随俗
鲨鱼和鲸鱼就是上文中第一种情况的代表。鲨鱼是软骨鱼纲、鲨总目下 500 多个物种的总称,它们的族谱可以追溯到距今 3 亿多年前的泥盆纪,是妥妥的海洋原住民。而哺乳纲的鲸类的祖先原本是海边的四足走兽,几千万年前决定下海生存,才逐渐演化成如今的海洋大家族。
自从开启了海洋生活,鲸鱼逐渐 “入乡随俗”:四肢特化为鳍,身体呈流线型,外型和鱼类越长越像,不过保留了哺乳动物恒温、用肺呼吸、胎生等特点。

鲸的进化过程
来源:左 coast.noaa.gov 右 www.redbubble.com
吃的像,长得也得像
在鲸鱼和鲨鱼之中,还有更典型的趋同进化的例子,那就是滤食性的鲸鱼和鲨鱼。
须鲸家族是鲸鱼中的滤食者,它们利用口中的鲸须来过滤水中的小鱼小虾。例如蓝鲸会张开大嘴冲进磷虾群中,将海水和食物一并吞入,舌头挤掉海水后,就可以大饱口福了。

蓝鲸的大嘴巴,来源:CGTN
而滤食性的鲨鱼包括鲸鲨、姥鲨和巨口鲨,它们通过鳃耙来过滤水中的小型生物。(绵绵的餐桌之前讲过啦,忘记的回去复习哦!)

左:座头鲸的鲸须,来源:Whale and Dolphin Conservation
右:“绵绵” 的鳃耙
俗话说"You are what you eat",为了有利于滤食的取食方式,这几种生物都演化出了一些相似的特征。比如它们都有着巨大的嘴:蓝鲸的大嘴能张开到接近 90°,还有可以展开的喉腹褶;鲸鲨的嘴巴可以张到 1.5 米宽,姥鲨的嘴超过一米;而我们的巨口鲨 “绵绵”,全身上下最宽的地方就是嘴巴。再比如,它们都有用于过滤的结构,比如须鲸口中的鲸须和巨口鲨鳃弓上的鳃耙。

鲸鲨的大嘴巴,来源:左 theallineed.com/右 ladbible.com

姥鲨的大嘴巴,来源:wikia.nocookie.net

“绵绵” 的大嘴巴 左:实体 右:模型
为什么它们都演化出了滤食的捕食方式呢?
生物演化史上,在大型掠食者繁盛一时的情况下,竞争饱和,总有一支别出心裁的物种为了逃避内卷,从而跨食物链层级地选择更小的食物链底层但数量庞大的小生物群作为饵食,以此来提高进食效率、减少能量消耗,从而发展出巨大的体型。
须鲸家族早期也以掠食为生,但后来它们发现了更稳定的食物来源 —— 个体微小、数以亿计的浮游动物和群游小鱼。用滤食的方式,只要在海水中张大嘴游动就可以获取丰厚的蛋白质,这可比打打杀杀的捕猎要悠闲多了。吃得多,又没有支撑身体的限制,自然就越长越大。而对于巨口鲨来讲,深海食物稀少,浮游生物和小型动物是珍贵的食物来源。
巨口鲨吃饭
大自然中的英雄所见略同 | 趋同演化
古人类演化 2025 年 08 月 25 日 21:42 上海
在物种演化这台连续剧中,“自然选择” 扮演着金牌策划的角色,在环境的舞台上,能让更适应的特征留下来。那么问题来了:如果不同的物种面对着相似的环境压力,自然选择这一 “金牌策划” 有没有可能做出相同或相似的筛选,从而使得不同物种被筛选出相似的特征?
其中的答案就藏在生物学中独特的现象:“趋同演化” 之中。趋同演化(Convergent Evolution) 指的是:一些亲缘关系并不密切的物种,在相似的环境压力下,分别独立地进化出了相似的形态或功能。换句话说,虽然它们的 “剧本来源” 不同,但因为舞台条件相似,最终呈现的角色特征却惊人地接近。趋同进化的思想可以追溯到 19 世纪的英国生物学家理查德・欧文对同源与类似的区分。尽管他对达尔文的进化论持怀疑态度,但在 19 世纪中期,他指出了一个关键区别:一些动物的身体部分在结构上相似(同源结构),而另一些则只是功能相似(类似的结构)[1]。
例如,海豚的鳍状肢与人类的手是同源结构,因为它们共享相同的骨骼框架,只是自上一个共同祖先以来,功能发生了分化。相比之下,海豚的鳍状肢与鲨鱼的鳍则属于类似结构 —— 它们在功能上相同,都是用来游泳的,但在形态和进化来源上却不同,因为这是各自独立、通过趋同进化形成的。
从下图可以看到:左边的海豚,右边的鲨鱼,它们是不是看起来很像?都有流线型的身体、背鳍和胸鳍,都是海洋里的 “游泳健将”。可你知道吗?它们的亲缘关系其实比你和海豚还要遥远!鲨鱼和海豚的关系,就好比 “远房亲戚中的远房亲戚”,虽然同属脊椎动物,但血缘差得非常远;反而是海豚和人类,虽然模样和生活方式大相径庭,却算得上 “近亲”,因为我们同属哺乳动物。要记住,鲨鱼是鱼类,而海豚是哺乳动物,和人类一样用肺呼吸、用母乳哺育下一代。

海豚和鲨鱼分别进化出了非常相似的外形特征,这些特征帮助它们成为成功的海洋捕食者。(图片来源:Shutterstock)
除此之外,在植物中,也能找到许多趋同演化的实例。以植物为例,美洲的仙人掌与非洲的大戟科植物虽 “家世” 迥异,却在干旱的沙漠环境中都演化出了肥厚的肉质茎和尖锐的刺,用以储存水分并减少蒸腾 [2]。随着分子生物学技术的进步,人们也逐渐开始探究这些过程是否在分子水平上可以观察到。例如美国密歇根州立大学 Robert VanBuren 实验室通过比较基因组和转录组学研究,揭示了草类植物在耐旱性方面的趋同进化机制,展示了环境压力如何在分子层面塑造相似的适应特征 [3]。
趋同演化之所以会出现,核心原因在于相似的环境压力会驱动物种演化出相似的适应策略。生物体在生存和繁衍过程中,总是要面对自然选择的考验。当不同的物种处于类似的环境或生态位时,即便它们的 “出身背景”(进化谱系)完全不同,也可能被 “逼迫” 朝着相似的方向进化。
环境选择压力(Environmental Selection Pressure)
例如,水生环境要求身体流线型、呼吸和运动高效,于是鲨鱼和海豚都进化出流线体型和鳍状结构。
功能上的约束(Functional Constraints)
某些功能只有有限的解决方案。比如沙漠植物储水、防蒸腾的有效方式就是 “长肉茎 + 减少气孔密度或改变气孔结构 + 刺”,所以无关的仙人掌和大戟科植物就不约而同地演化出了类似形态。
进化的 “工具箱” 有限(Limited Evolutionary Toolkit)
所有生物的基因和发育途径都是有限的,面对同样的问题,只能在这些 “工具” 中找到可能的解决办法,因此更容易出现相似的进化结果。
换句话说,趋同演化就像是不同选手参加同一场比赛:起点不同、背景各异,但规则和目标相同,最后往往会跑出相似的路线。所以,下次当你看到两种长得很像但八竿子打不着的生物时,就可以大胆猜测:“它们是不是经历了趋同演化?” 这背后真正的导演,就是 “自然选择”。是生存的压力,让生命在不同的演化道路上,不约而同地走向了相似的辉煌。这就是大自然的 “英雄所见略同”!
参考文献
1.Owen, R., and Cooper, W.W. (1843). Lectures on the Comparative Anatomy and Physiology of the Invertebrate Animals: Delivered at the Royal College of Surgeons, in 1843 (Longman, Brown, Green, and Longmans).
2.Bennici, A. (2003). The convergent evolution in plants. Riv Biol 96, 485-489.
3.Marks, R.A., Van Der Pas, L., Schuster, J., Gilman, I.S., and VanBuren, R. (2024). Convergent evolution of desiccation tolerance in grasses. Nat Plants 10, 1112-1125. 10.1038/s41477-024-01729-5.
狗狗:出走的决心
原创 莫喻枫 赛先生 2025 年 08 月 26 日 19:24 广东

一只澳洲野狗站在岩石上,目光凝视远方,神情警惕而担忧。它竖起耳朵,眉头紧锁。它的皮毛呈褐色,腹部和面部区域有白色的毛。图片:© 澳大利亚博物馆
导读:
国际狗狗日(International Dog Day)是美国动物保护倡议者 Colleen Paige 发起的一个节日,定在每年的 8 月 26 日,以关注狗在社会中的角色与福利。实际上,狗作为人类最早驯化的动物之一,与人类相伴已有三万余年,并走过了与人类协同进化、出走野外的精彩旅程。
莫喻枫 | 撰文
在人类与动物共同演化的漫长历史中,狗演绎着一段独特的 “双重进化” 传奇。作为最早被人类驯化的伙伴,它们从狼群中走出,历经万年的自然与人工选择,演化成形貌各异的家犬,深度融入人类社会的每一个角落。
然而,在历史的长河中,也有一批犬群脱离人类,出走荒野,在自然选择的塑造下开启了一场逆向的旅程 —— 从东亚家犬野化为澳洲野犬。在这个过程中,它们重塑了行为、代谢和神经机制,重新适应野外生存。
驯化与野化,两条相反的演化之路,如何在狗狗的基因组上留下各自的烙印?
狗狗与人类走上相似的基因进化之路
中国科学院昆明动物研究所、动物进化与遗传前沿交叉卓越创新中心的张亚平团队长期致力于研究家养动物的起源和驯化过程。在一系列的工作中,他们对多种灰狼、中国本土犬以及其他现代犬种进行了全基因组测序和对比分析 [1-4]。
通过分析这些数据,他们发现,狼和中国本土犬大约在 3.2 万年前开始分化。最早的狗狗可能是在靠近人类营地、一起觅食的过程中,逐渐走上驯化之路的。
在狗的驯化过程中,有一批基因受到了 “正向选择”—— 也就是更有利于适应人类环境的基因被保留了下来。有趣的是,这些基因与人类在自身进化过程中被选择的基因有很大重叠,尤其在消化代谢、大脑神经功能以及癌症相关基因方面,表现出明显的 “平行进化”—— 即人和狗在相似的环境压力下,演化出了类似的基因改变。
从狼到狗的驯化历程伴随着饮食结构的深刻转变。约一万年前,人类祖先主要依赖狩猎和采集为生。随着农业的兴起,大多数人群逐渐从狩猎 - 采集的生活方式转向定居的农业社会。在这一重大转型过程中,人类与狗共同经历了对富含淀粉饮食的适应。
以高淀粉食物为主的人群,其唾液淀粉酶基因 AMY1 的拷贝数更多,能够表达更高水平的淀粉酶,从而更有效地消化淀粉类食物。值得注意的是,狗也演化出了类似的适应性特征。在农业起源较早的地区,犬类的胰腺淀粉酶基因 AMY2B 拷贝数显著增加,基因表达水平也随之升高,增强了它们对淀粉的消化能力 [2]。
另一个基因改变的有力证据来自高原适应。藏獒最初起源于中国平原地区的本土犬,后来逐渐进入青藏高原并完成了对极端环境的适应。研究人员比较了 32 只藏獒、20 只中国本土犬和 14 只灰狼的全基因组单核苷酸多态性(SNP,是基因组中最常见的遗传变异类型)分型,共识别出超过 12 万个 SNPs,并进一步筛选出在藏獒中经历正向选择的 16 个基因,其中有 12 个与高海拔低氧适应密切相关,例如 EPAS1、SIRT7、PLXNA4 和 MAFG [3]。
其中,一种缺氧诱导因子(HIF)—— EPAS1,同样在藏族人群的低氧适应中发挥作用 [5],这支持了狗与人类在高海拔适应方面可能存在趋同进化的可能性。
在人与犬类共同 “正向选择” 的基因中,与大脑和神经活动相关的基因尤其值得关注。其中一个关键基因是 SLC6A4,它负责调节大脑中血清素的运输,而血清素是影响情绪、行为和社交能力的重要神经递质。驯化往往意味着生活环境拥挤,这些 “不太友好” 的条件会对基因产生压力,推动基因组发生改变。比如,神经系统通路中的一些基因,尤其是与血清素相关的基因,可能因为能降低攻击性而被保留下来,以便更好适应群体生活 [1]。
有意思的是,那些在人类和狗身上被 “选中” 的基因,虽然帮助我们更好地适应了共同的生活环境,却也可能带来了相似的健康问题。比如,一些有助于缓解压力、增强社交能力的基因变化,可能同时增加了患某些神经系统疾病的风险。此前,张亚平团队曾报道犬类强迫症(Canine compulsive disorder, CCD)与人类强迫症(Obsessive-compulsive disorder, OCD)之间有多个共享的风险基因 [6],在遗传构成上存在强烈的趋同性。
正因为人类和狗在相似的环境下经历了类似的基因演化,狗可能成为研究人类疾病的重要 “参照”。
当东亚家犬出走澳洲,重返野外
一方面,狗在与人类相处的过程中被人类驯化,另一方面,一些被驯化的狗狗在机缘巧合之下又重返野外,走出了另外一条演化道路。
例如,数千年前,一群源自东亚的家犬随人类抵达澳大利亚。在这片陌生大陆上,它们逐渐脱离人类的驯养,回归荒野,开启了独立狩猎、自主繁衍的生存模式,并逐步适应了当地的自然环境。
历经数千年的野化,这群犬类最终演化为今天的澳洲野犬(Canis dingo)。
尽管野化现象备受关注,但其背后的基因组机制仍知之甚少。主要难点在于,多数野化种群常与野生或家养祖先发生基因交流,难以区分遗传变化是源于自然选择,还是杂交带来的基因流动。
而澳洲野犬正是破解这一难题的理想模型。其野化进程始于数千年前,并且直至近 200 年欧洲人到来之前,一直与家犬及狼等祖先种群保持地理与遗传上的隔离。这一独特的演化历史,有效避免了杂交干扰。
为了进一步理解这一过程,张亚平团队通过对澳洲野犬及其相关犬科动物进行全基因组测序,并整合分析多个已发表的犬科动物全基因组数据,详细揭示了澳洲野犬的起源与演化历程。
研究证实,澳洲野犬的祖先是东亚地区已被驯化的家犬,大约 9900 年前从中国南方出发,于约 8300 年前抵达澳大利亚,并在当地野化 [7-8]。
值得注意的是,这一时间早于著名的 “南岛语族扩散” 约两千年。南岛语族约五六千年前从福建一带经台湾向太平洋和印度洋岛屿迁徙。澳洲野犬的更早到来,暗示可能存在一次尚未被充分认知的古代人类迁徙活动,将家犬带入澳洲 [9]。
研究人员在澳洲野犬处于正向选择压力下的基因组区域中鉴定出了 50 个候选基因。正在经历 “正向选择” 的基因多与消化、代谢、繁殖和神经发育相关。例如,与消化相关的 SLC5A1 基因在葡萄糖和钠的吸收中起重要作用;TAS2R5 基因可能参与苦味感知;ARHGEF7 基因可能促进海马神经元发育;Prss37 基因与繁殖相关,在小鼠中被证明对雄性生育能力至关重要 [10]。
正如前文所提及,家犬与野生犬科动物如狼和澳洲野犬之间存在着显著的饮食差异。野生犬科动物主要以肉类为食,而家犬通常会摄入相当数量的人类提供的植物食物。家犬淀粉消化能力的增强是通过胰腺淀粉酶 (AMY2B) 基因拷贝数的扩增实现的,但澳洲野犬拥有与狼相同的未扩增基因拷贝数。
与澳洲野犬生育力相关基因被 “正向选择” 相似,此前的野化鸡和野化水稻的研究也显示,生殖机制的改变是动植物野化过程中的一个普遍且关键的适应性特征 [11-12]。在野化水稻中,研究者鉴定出的与新环境适应相关的基因组区域主要与开花时间、繁殖特性及应激反应相关。在野化鸡中,则显著地识别出与性选择和繁殖相关的基因可能构成了推动野化进程的关键自然选择靶点。这表明,无论是动物还是植物,适应新环境往往伴随着饮食与繁殖策略的双重调整。
同时,狗的野化过程对与神经发展相关的基因组区域产生了正向选择。其中两个关键基因是 ARHGEF7 和 PANX2。 ARHGEF7 基因可促进海马神经元树突棘和突触的形成。海马体在反应抑制、记忆和空间认知中发挥关键作用,部分研究提示其与目的性行为相关。因此,该基因表达水平的改变可能与澳洲野犬适应野外生存环境的行为变化有关。
结语
狗狗在驯化与野化过程中的基因演变,生动展现了自然环境与人类社会文化如何共同塑造生命体的基因组。
对科学而言,狗是研究复杂性状遗传、疾病机制与衰老规律的天然模型;对人类而言,狗不仅是最早的伙伴,更是理解我们自身进化历程的一面镜子。探索犬类的驯化与野化,让我们在基因与历史的交汇之处,重新审视人与动物、文明与自然之间那段深远而深刻的关系。
##参考文献:
[1] Wang, Guo-dong, et al. “The genomics of selection in dogs and the parallel evolution between dogs and humans.” Nature communications 4.1 (2013): 1860.
[2] Wang, Guo-Dong, et al. “Structural variation during dog domestication: insights from gray wolf and dhole genomes.” National Science Review 6.1 (2019): 110-122.
[3] Li, Yan, et al. “Population variation revealed high-altitude adaptation of Tibetan mastiffs.” Molecular biology and evolution 31.5 (2014): 1200-1205.
[4] Li, Yan, et al. “Artificial selection on brain-expressed genes during the domestication of dog.” Molecular biology and evolution 30.8 (2013): 1867-1876.
[5] http://www.kiz.cas.cn/gre/gre7/gre73/201408/t20140827_4192627.html
[6] Cao, Xue, et al. “Whole genome analyses reveal significant convergence in obsessive-compulsive disorder between humans and dogs.” Science Bulletin 66.2 (2021): 187-196.
[7] Zhang, Shao-jie, et al. “Genomic regions under selection in the feralization of the dingoes.” Nature Communications 11.1 (2020): 671.
[8] https://www.cas.cn/syky/202003/t20200302_4736163.shtml
[9] http://env.people.com.cn/n1/2020/0317/c1010-31635368.html
[10] Shen, Chunling, et al. “Prss37 is required for male fertility in the mouse.” Biology of reproduction 88.5 (2013): 123-1.
[11] Qiu, Jie, et al. “Genomic variation associated with local adaptation of weedy rice during de-domestication.” Nature communications 8.1 (2017): 15323.
[12] Johnsson, Martin, et al. “Feralisation targets different genomic loci to domestication in the chicken.” Nature communications 7.1 (2016): 12950.
via:
-
Convergent Evolution - Definition and Examples | Biology Dictionary
https://biologydictionary.net/convergent-evolution/ -
原本不一样,现在长得像:趋同进化是个啥?
https://mp.weixin.qq.com/s/u8guCLlKWTYd4okISxpSeA -
大自然中的英雄所见略同 | 趋同演化
https://mp.weixin.qq.com/s/t_LbXgDb7bzf6A3UNGlVSA

















 949
949

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









