







多组学分析揭示甘蓝型油菜的 AGR-FC.C3 基因座是控制花瓣颜色的新候选者
Abstract
Variations in the petal color of Brassica napus are crucial for ornamental value, but the controlled loci for breeding remain to be unraveled. Here, we report a candidate locus, AGR-FC.C3, having conducted a bulked segregant analysis on a segregating population with different petal colors. Our results showed that the locus covers 9.46 Mb of the genome, harboring 951 genes. BnaC03.MYB4, BnaC03.MYB85, BnaC03.MYB73, BnaC03.MYB98, and BnaC03.MYB102 belonging to MYB TFs families that might regulate the petal color were observed. Next, a bulk RNA sequencing of white and orange-yellow petals on three development stages was performed to further identify the possible governed genes. The results revealed a total of 51 genes by overlapping the transcriptome data and the bulked segregant analysis data, and it was found that the expression of BnaC03.CCD4 was significantly up-regulated in the white petals at three development stages. Then, several novel candidate genes such as BnaC03.ENDO3, BnaC03.T22F8.180, BnaC03.F15C21.8, BnaC03.Q8GSI6, BnaC03.LSD1, BnaC03.MAP1Da, BnaC03.MAP1Db, and BnaC03G0739700ZS putative to controlling the petal color were identified through deeper analysis. Furthermo re, we have developed two molecular markers for the reported functional gene BnaC03.CCD4 to discriminate the white and orange-yellow petal colors. Our results provided a novel locus for breeding rapeseed with multi-color petals.
油菜(Brassica napus)花瓣颜色的变异对其观赏价值至关重要,但控制该性状的基因座尚未完全揭示。本研究通过对具有不同花瓣颜色的分离群体进行集群分离分析,报告了一个候选基因座AGR-FC.C3。结果表明,该基因座覆盖了9.46 Mb的基因组区域,包含951个基因。我们观察到MYB转录因子家族中的BnaC03.MYB4、BnaC03.MYB85、BnaC03.MYB73、BnaC03.MYB98和BnaC03.MYB102等基因,可能调控花瓣颜色。接着,我们对白色和橙黄色花瓣在三个发育阶段进行了集群RNA测序,以进一步识别可能受调控的基因。结果通过重叠转录组数据与集群分离分析数据,识别出51个基因,并发现BnaC03.CCD4基因在三个发育阶段的白色花瓣中显著上调。随后,通过更深入的分析,鉴定出若干新的候选基因,如BnaC03.ENDO3、BnaC03.T22F8.180、BnaC03.F15C21.8、BnaC03.Q8GSI6、BnaC03.LSD1、BnaC03.MAP1Da、BnaC03.MAP1Db和BnaC03G0739700ZS,这些基因可能与花瓣颜色的控制有关。进一步地,我们为已知功能基因BnaC03.CCD4开发了两个分子标记,用于区分白色和橙黄色花瓣。我们的研究为多色花瓣油菜的育种提供了一个新的基因座。
1. Introduction
B. napus is an important oilseed crop, supplying a primary source of edible vegetable oils and high-protein animal feeds [1,2,3]. The petal color is an essential indicator of agronomic traits in rapeseed, and flowers with rich colors have played a vital role in attracting insects and other pollinators for adequate pollination [4,5]. Additionally, the petal color in B. napus also holds ornamental value, and economically beneficial flowers with specific colors have been obtained [6,7,8,9].
油菜(B. napus)是一种重要的油料作物,是食用植物油和高蛋白动物饲料的主要来源[1,2,3]。花瓣颜色是油菜农艺性状的重要指标,色彩丰富的花朵在吸引昆虫和其他传粉者进行充分授粉方面发挥了重要作用[4,5]。此外,油菜的花瓣颜色也具有观赏价值,通过选育具有特定颜色的花朵,可以获得具有经济价值的花卉[6,7,8,9]。
Flower coloration is primarily determined by the content of carotenoids, flavonoids, and betalains [4]. Carotenoids are responsible for developing yellow, orange, and red flower colors [4,10]. The enzymes responsible for cleaving specific double-bond regions of carotenoids are known as carotenoid cleavage oxygenases (CCOs) or carotenoid-cleaving dioxygenases (CCDs) [5]. For instance, the CmCCD4a gene is expressed explicitly in chrysanthemum petals, degrading carotenoids into colorless compounds and changing petal color from yellow to white [11]. Similarly, the flesh was yellow due to carotenoid accumulation in the PpCCD4 mutant of peaches [12]. Mutations in the CrtR-b gene, which explicitly expresses β-carotene hydroxylase in tomato flowers, significantly decrease carotenoid levels and result in white petals [13]. The carotenoid isomerase (CRTISO) is crucial in carotenoid synthesis by converting the prolycopene to all-trans lycopene [14]; it was found that the pro-lycopene was accumulated in the mutation of CmCRTISO and led to the petal color changing from yellow to light yellow in melon [15]. The function of BrCRTISO1 has also been confirmed to control the yellowish color of petals in B. rapa [16]. A significant reduction in total carotenoid content resulted in a creamy-white phenotype in petals through the site-directed mutagenesis of BnaCRTISO in B. napus [17].
花瓣颜色主要由类胡萝卜素、黄酮和甜菜碱的含量决定[4]。类胡萝卜素负责形成黄色、橙色和红色的花朵颜色[4,10]。负责裂解类胡萝卜素特定双键区域的酶被称为类胡萝卜素裂解氧化酶(CCOs)或类胡萝卜素裂解二氧化酶(CCDs)[5]。例如,CmCCD4a基因在菊花花瓣中显著表达,能够降解类胡萝卜素为无色化合物,并将花瓣颜色从黄色改变为白色[11]。类似地,在桃子中的PpCCD4突变体由于类胡萝卜素的积累使果肉呈黄色[12]。番茄花朵中显著表达β-胡萝卜素羟化酶的CrtR-b基因的突变会显著降低类胡萝卜素的含量,并导致花瓣变白[13]。类胡萝卜素异构酶(CRTISO)在类胡萝卜素合成中起着至关重要的作用,通过将前番茄红素转化为全反式番茄红素[14];在甜瓜的CmCRTISO突变体中发现前番茄红素积累,导致花瓣颜色从黄色变为浅黄色[15]。BrCRTISO1的功能也已被证实能控制油菜(B. rapa)花瓣的黄色[16]。通过对油菜(B. napus)中的BnaCRTISO进行定点诱变,类胡萝卜素总含量显著降低,导致花瓣呈奶油白色表型[17]。
Flavonoids play an important role in forming petal color [18]. Anthocyanins (a subclass of flavonoids) are widespread in vascular plants and are responsible for various colors in flowers, such as orange, red, purple, and blue [4]. Manipulating or modifying genes that are associated with anthocyanin synthesis or its precursors has been successfully used in altering the petal color in several plant species, for example, in Torenia hybrida, Petunia hybrida, Nicotiana tabacum, and Eustoma grandiflorum (Raf.) Shinn [19,20,21,22,23]. In red-flowered rapeseed, various colors ranging from raspberry red to beige red and zinc yellow can be achieved via RNA interference to target the anthocyanidin synthase (ANS) BnaA03.ANS gene at different levels [24].
类黄酮在形成花瓣颜色中发挥重要作用[18]。花青素(类黄酮的一个亚类)广泛存在于维管植物中,负责花朵中多种颜色的呈现,如橙色、红色、紫色和蓝色[4]。通过操控或修改与花青素合成及其前体相关的基因,已经成功地改变了多种植物物种的花瓣颜色,例如在紫茉莉(Torenia hybrida)、矮牵牛(Petunia hybrida)、烟草(Nicotiana tabacum)和大丽花(Eustoma grandiflorum (Raf.) Shinn)中[19,20,21,22,23]。在红色花朵油菜中,通过RNA干扰不同程度地靶向花青素合成酶(ANS)BnaA03.ANS基因,可以实现从覆盆子红到米色红和锌黄等多种颜色[24]。
Other regulation factors have also been uncovered to influence petal color. In B. juncea, the stable storage of carotenoids was disrupted in the mutation of BjA02.PC1 and BjB04.PC2 (belonging to the esterase/lipase/thioesterase family of acyltransferases) led to a white-flowering phenotype [25]. Additionally, a specific rapeseed line with a pair of Orychophragmus violaceus chromosome M4 exhibited reddish petals [26], and further analysis showed that the OvPAP2 transcript of O. violaceus is expressed specifically in rapeseed petals [27].
其他调控因素也已被发现对花瓣颜色产生影响。在芥菜(B. juncea)中,BjA02.PC1和BjB04.PC2基因的突变(属于酯酶/脂肪酶/硫酯酶家族的酰基转移酶)导致类胡萝卜素的稳定储存受到干扰,从而表现出白色花瓣的表型[25]。此外,一种特定的油菜品种与紫花忍冬(Orychophragmus violaceus)染色体M4对偶体表现出红色花瓣[26],进一步分析表明,紫花忍冬的OvPAP2转录本在油菜花瓣中有特异性表达[27]。
Bulked segregant analysis (BSA) is a commonly used method that identifies the molecular markers associated with a specific gene or genomic region [28]. For example, CCD4, which had functions for controlling the petal color, was identified by combining BSA and positional cloning in B. napus [29]. Additionally, a transcriptomic and metabolomic analysis revealed that the reduced lutein and zeaxanthin are the primary reason for the appearance of white-flowered rapeseed [29], and the related genes (BnaA09.ZEP and BnaC09.ZEP) that encoded zeaxanthin epoxidase (ZEP) were identified using BSA in B. napus [30].
大规模分离群体分析(BSA)是一种常用的方法,用于识别与特定基因或基因组区域相关的分子标记[28]。例如,控制花瓣颜色的CCD4基因通过结合BSA和定位克隆在油菜(B. napus)中被鉴定出来[29]。此外,转录组学和代谢组学分析揭示,叶黄素和玉米黄质的减少是白花油菜出现的主要原因[29],并且通过BSA在油菜中鉴定了与之相关的基因(BnaA09.ZEP和BnaC09.ZEP),这些基因编码玉米黄质环氧化酶(ZEP)[30]。
The molecular mechanisms underlying flower coloration are still not fully understood. In this study, F1 was first obtained by hybridizing the B. napus with orange-yellow petals (YP) and white petals (WP). Then, an F2 segregation population with a series of petal colors was obtained. The extreme material pools with WP and YP in F2 were selected for BSA-Seq and RNA-Seq analyses, and the associated genomic region (AGR) of AGR-FC.C3 that might control the petal colors and some key genes involved in carotenoid pathways were identified. These findings could enhance the breeding process of ornamental rapeseed and provide a theoretical foundation for future breeding with white and orange-yellow flowers in B. napus.
花朵着色的分子机制仍未完全明了。在本研究中,首先通过杂交具有橙黄色花瓣(YP)和白色花瓣(WP)的油菜(B. napus)获得了F1代。随后,获得了具有一系列花瓣颜色的F2分离群体。选择了F2代中具有极端白色花瓣(WP)和橙黄色花瓣(YP)的材料池进行BSA-Seq和RNA-Seq分析,鉴定出了可能控制花瓣颜色的基因组相关区域(AGR)以及一些与类胡萝卜素通路相关的关键基因。这些发现有助于提高观赏油菜的育种过程,并为未来白色和橙黄色花朵油菜的育种提供了理论基础。
2. Results
2.1. BSA for Petal Colors in B. napus
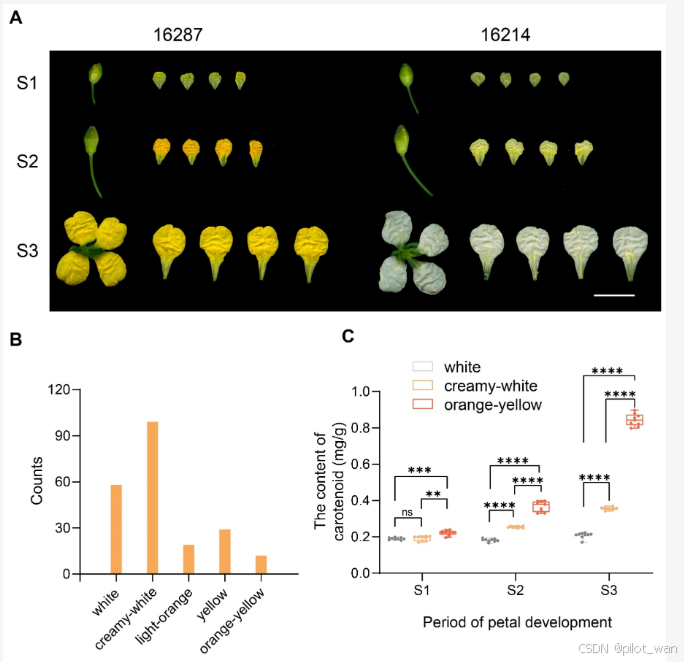
The reciprocal hybridization between B. napus 16214 and 16287 was performed (Figure 1A). It was revealed that all F1 plants, no matter 16214 × 16287 or 16287 × 16214, had light-orange petals. The F1 seeds were subsequently sown in a field to yield the F2 generation. Further observation revealed that five distinct petal colors (white, creamy-white, light-orange, yellow, and orange-yellow) existed in the F2 generation (Figure 1B). Further analysis showed that the content of carotenoid in YP was significantly higher than in WP, particularly in S3, which was approximately three-fold more elevated than that of the WP (Figure 1C).
对油菜品种16214和16287进行了互交杂交(图1A)。结果显示,无论是16214 × 16287还是16287 × 16214,所有F1植株的花瓣均为浅橙色。随后将F1种子播种在田间,获得了F2代。进一步观察发现,F2代中存在五种不同的花瓣颜色(白色、奶白色、浅橙色、黄色和橙黄色)(图1B)。进一步分析显示,橙黄色(YP)花瓣中的类胡萝卜素含量显著高于白色(WP)花瓣,特别是在S3阶段,其含量大约是白色花瓣的三倍(图1C)。
图1A图1B图1C 图1. 油菜橙黄色和白色花朵的表型变异。
(A) 16287和16214在S1、S2和S3阶段的表型。条形图,1 cm。
(B) F1代中不同花瓣颜色的个体数量。
(C) 类胡萝卜素含量的测定。三次重复实验,误差条表示标准差。每个样本有九个生物学重复。** p < 0.01, *** p < 0.001, **** p < 0.0001;ns,表示无显著差异。
To investigate the genetic basis underlying the color difference, BSA was conducted on extreme pools with WP and YP in F2 lines, respectively. A total of 147.32 GB of clean data was obtained with a Q30 of 94.48% (Table S1). Each sample showed an average alignment rate of 93.50% to the reference genome, with an average 28× coverage depth and genome coverage of 94.90%, which satisfied the subsequent analysis requirements (Table S2). Genome-wide, a total of 668,993 single nucleotide polymorphisms (SNPs) and 668,993 insertion–deletions (InDels) between 16214 and 16287 were identified (Figure 2, Tables S3 and S4). It was revealed that a significant genomic region of 9.46 Mb (63.03–72.49 Mb) on ChrC03 was associated with petal color (Figure 2 and Figure 3A). Since the Darmor_bzh genome has a gap in this region, this region in the Darmor_bzh was also mapped to the genome of Zhongshuang 11 (ZS11) (Figure 3B). Further analysis showed that a significantly associated regi
 最低0.47元/天 解锁文章
最低0.47元/天 解锁文章



















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









