



利用亚特兰蒂斯生态系统模型探索北欧和巴伦支海的平衡捕捞
摘要
通过设置不同捕捞压力以及按计算出的生产力成比例实施平衡捕捞模式的情景,我们研究了在生态系统中捕捞不同组分的交互效应。这些模拟得出的结论是:
– 模型结果表明,在当前挪威海和巴伦支海实施BH制度将带来更高的总产量,主要来自较低营养级的物种。
– 挪威渔业受到良好监控和管理,在平衡捕捞模式方面已经得分较高,大多数物种的捕捞水平接近各自的最大可持续产量(MSY),这表明在当前商业种群上实施BH制度带来的收益有限。
然而,结果证实,将渔业扩展到目前未进行商业收获的物种仍有空间,尽管需要强调的是,这可以在不全面实施平衡收获的情况下实现,前提是能够找到商业市场。
我们应该承认,当前模型中非商业物种的适当捕捞水平和模型参数化需要进一步验证,因为这些非商业物种出人意料地比商业物种更容易崩溃,这在逻辑上并不合理。
当前模型还发现了其他一些缺陷和难以解释的结果,这说明构建稳健的端到端模型需要大量的综合数据。此外,生物产量数据目前并未作为亚特兰蒂斯模型的输出内容,而本研究中用于估算生产力水平的方法也尚未被尝试或测试过。
这是首次使用大西洲模型对水生生态系统的大部分生物组分进行平衡捕捞模拟,所有结果仅应视为初步结论。通常,在探索未知且未经检验的领域时,我们最终提出的问题多于答案。然而,结果的大致轮廓可能既稳健又具有普遍性。我们的研究结果表明,尽管将BH与过度捕捞的生态系统相比可能显示出存在潜在收益,但将BH与相对管理良好的生态系统相比则呈现出更为复杂的局面。对于当前已被开发的物种而言,相较于现有的相对管理良好的渔业制度,采用BH模式带来的经济效益极为有限;尽管高度选择性的渔业所造成的不利的生物副作用可能从BH制度中获益更多。大多数已识别的收益来源于降低对过度捕捞物种的捕捞压力,并将开发扩展到目前未捕捞或捕捞程度较轻的物种。
尽管BH提倡对“生态系统的各种物种、种群和体型大小进行尽可能广泛的捕捞”(加西亚等,2012),但应注意其中的“可能”一词。BH并不要求必须应用于生态系统中的每一个组成部分。例如,即使在BH模式下,社会也可能选择将旗舰性大型动物排除在捕捞范围之外;或者某些物种和体型类别可能因捕捞成本过高而不具经济性。因此,在评估BH可能产生的影响时,有必要采用具有足够水平的模型在细节方面(例如亚特兰蒂斯),以检验平衡收获不同潜在实施方案的具体结果。我们希望本文已经证明,这种程度的分析现在是可以实现的。
最后需要指出的是,随着人类人口快速增长,到2050年可能接近90亿(联合国,2019),健康食品的需求已成为我们世界面临的一大挑战。联合国可持续发展目标(SDGs)将可持续发展目标2中的零饥饿目标、可持续发展目标3中的良好健康,以及可持续发展目标14中的保护和可持续利用水下生物列为全球17个最重要问题中的三项(联合国,2015)。对营养和健康食品的需求从未如此重要,迫切需要制定新的可持续捕捞策略,以确保增加食物生产的同时不耗尽生态系统。目前,仅有3%的食物来自海洋(菲尔德等人,1998年),这表明潜力尚未被开发。本研究展示了在不对系统造成重大干扰的情况下,将我们的收获模式扩展到未开发物种的潜力,这也是平衡收获的主要目标之一。
引言
如今,渔业普遍被认为处于资源匮乏的状态,几乎没有进一步扩展的空间,甚至有人宣称,如果当前趋势持续下去,未来50年内将无鱼可捕(Black, 2006)。根据联合国粮农组织的统计数据,全球海洋捕捞渔业产量三十多年来一直持平,且被认定为过度捕捞的未评估种群数量日益增加,被视为过度捕捞(FAO, 2018)。现代渔业管理面临的主要挑战之一是制定捕捞策略,在确保高效和最大可持续利用海洋生产力(《联合国海洋法公约》, 1982)的同时,保护捕捞种群及生态系统的结构和功能(《生物多样性公约》, 1992)。基于1998年马拉维原则(联合国环境规划署/《生物多样性公约》, 1998)提出的渔业生态系统方法(EAF)概念,已被建议作为应对这些目标的整体性框架。
挪威依法承诺在北海、挪威海和巴伦支海实施基于生态系统的渔业管理(EBFM)(Miljøverndepartementet, 2006, 2009, 2011;Olsen et al., 2007)。根据Pitcher et al. (2009)的观点,按照负责任渔业行为守则(FAO, 1995),在挪威实施符合生态系统原则的管理应相对简单。尽管目前仍主要受传统单物种管理规范的约束,但挪威在基于生态系统的原则方面已处于较高水平。一个考虑生态系统的例子是东北北极鳕鱼和巴伦支海毛鳞鱼的管理,自1991年以来,在毛鳞鱼渔业中已考虑到毛鳞鱼作为鳕鱼食物的重要性(ICES, 2015a)。尽管EBFM的基本原则已达成共识,但在具体实施方面仍存在不确定性,例如如何在“开发”与“保护”之间取得平衡(Howell et al., 2016)。
为了实现渔业调整框架(EAF)的目标,“平衡捕捞”(以下简称BH)被提议为一种可能的策略,以确保获得较高的可持续产量,同时维持生态系统结构与功能。加西亚等人(2012)将BH定义为“在整个生态系统的尽可能广泛的物种、种群和体型大小范围内,根据其自然生产力施加适度的捕捞压力,从而保持其相对大小和物种组成。” 显然,BH并不是必要组成部分。它也不能成为完整的解决方案,因为任何总体捕捞策略都需要与其他策略协同实施,例如保护脆弱生境的策略。然而,BH已被提议作为完整的基于生态系统的渔业管理的一个可能组成部分,因此我们试图分析在特定生态系统中实施BH可能带来的后果。
需要注意的是,平衡收获(BH)在科学文献中一直存在相当大的争议,有支持也有反对这一概念的论文。平衡收获(BH)的概念在伦理和理论依据上受到了诸多批评(例如,伯吉斯等人,2016;弗罗泽等人,2016;保伊等人,2016)。其中一个问题是可行性:在生态系统的广泛范围内进行捕捞在多大程度上是可行的(例如,豪厄尔等人,2016)。这一反对意见包括捕捞某些生态系统组分的实际困难、提供支持此类捕捞的科学建议的难度,以及此类捕捞的经济性,可能导致部分渔业无法盈利。
我们提出的问题是:“在假设模型具有最佳真实性的前提下,对于一个已经得到相对良好管理的生态系统,如果在挪威和巴伦支海生态系统中实施平衡收获(BH)会发生什么?” (Figure 1)。具体而言,是否能够获得收益,如果可以的话,它们来自哪里,以及系统相应的损失或结构变化是什么?
我们明确不涉及此类渔业的可行性问题,也不尝试对这种变化的经济或社会经济进行建模。这些问题很有价值,但超出了当前工作的范围。
平衡捕捞可被视为一种可能的方法,通过使生态系统中尽可能多的组分承受与其特定生产力成比例的捕捞死亡率,从而将渔业管理提升到生态系统层面。这一理念已引起全球广泛关注,并得到了非洲湖泊生态系统中小规模渔业实证研究(科尔丁和范齐滕,2014;科尔丁等,2015)以及海洋系统模型研究(加西亚等,2012;劳等,2013)的支持。这些研究表明,与当前的选择性捕捞相比,平衡捕捞可能在维持生态系统结构的同时提高总可持续渔获量。
平衡捕捞(BH)的概念源于对传统选择性捕鱼管理所引发问题的普遍担忧,这些问题包括全球渔获量停滞(联合国粮农组织,2016)、目标物种过度捕捞(科斯特洛等,2012;苏迈亚等,2012)、大型掠食性鱼类枯竭(克里斯滕森等,2014)以及可能导致早熟、进而产生更小体型鱼类的渔业诱导进化(海诺和戈多,2002;劳,2007;谢等,2010)。选择性捕捞在历史上根深蒂固,渔民通常出于经济和伦理原因(科尔丁和范齐滕,2011)而针对最大个体和物种进行捕捞。然而,任何形式的选择性移除都不可避免地会改变种群组成,从而影响生态系统结构和生物多样性——即使在中等捕捞强度下也是如此(加西亚等,2012)。
需要强调的是,平衡收获(BH)并非主张无选择性和无差别的捕鱼。事实上,有观点认为,平衡收获可能实际上需要更高水平的选择性(里德等人,2016)。平衡收获只是建议在生态系统层面采用一种不同类型的选择性,即总体捕捞压力根据生产力分布在不同物种和体型大小上,以维持生态系统结构(Garcia et al., 2015)。如果平衡收获能够使捕捞死亡率模仿类似捕食引起的自然死亡率,则对生活史特征的进化选择预期会降低。实施平衡收获将导致捕鱼船队更加多样化,使用更广泛的渔具,从而降低对任何特定性状产生渔业诱导选择的风险(周等,2019)。
由于生产力往往随体型大小的增加而降低(彼得斯,1986),向全面实施BH转变意味着减少对大型鱼类的捕捞,增加对小型物种和个体的捕鱼,而这些通常在工业国家被视为低价值且无法利用。尽管研究显示,BH在非洲小规模生计渔业中能够实现高生物量产量,并对生态系统体型谱产生较低影响,但尚不清楚这些结果是否适用于大规模现代商业海洋渔业(伯吉斯等人,2016;豪厄尔等人,2016)。
因此,有观点认为,任何BH的实施都将是部分实施(例如,豪厄尔等人,2016),我们试图通过运行模拟来区分其影响。
不同系统部分的BH。关于生态系统的哪些部分应被视为可收获资源也引发了伦理问题(例如,保伊等人,2016)。
最后,人们对所采用的建模技术提出了批评。通常(尽管并非完全如此),这些模型研究使用了简化的基于体型的模型结构,未能很好地解析生态系统组分的物种特异性动态。如上所述,我们并不打算在此介入这一理论争论。我们仅旨在探讨如果在巴伦支海的大西洲模型中实施BH可能会发生什么情况,并希望我们的研究结果能为整体讨论提供一些更具体的依据。本文不再进一步关注这一讨论,相关内容已在周等人(2019)的最新综述中涵盖,仅需指出的是,通过使用大西洲模型,我们力求在分析中尽可能包含较高的物种真实性。对于未能充分捕捉物种特异性细节的情况,我们将在讨论中予以说明。
目前,挪威渔业被认为管理得相当良好,大多数商业鱼类种群均采用捕捞控制规则(HCRs)进行收获,且捕捞压力适中。这些规则中的捕捞压力接近产生最大长期产量的水平,同时不会对种群造成过度捕捞的不必要风险,即接近国际海洋考察理事会(ICES)渔业管理中所使用的最大可持续产量(MSY)(ICES, 2018a)。挪威渔业在平衡收获原则(BH原则)方面表现突出,该原则主张针对不同营养级的一系列物种进行开发,其中包括对低营养级物种如哲水蚤属的挪威拟哲水蚤 Calanus finmarchicus (Calanus, 2018)以及高营养级物种如鸟蛋、海豹和鲸鱼的开发(Howell et al., 2016)。
然而,一些相对丰富的种群要么仅被轻微捕捞(例如极地鳕, Boreogadus saida ),要么完全未开发(例如中层鱼类)(国际海洋考察理事会,2016)。对于所有渔业,均适用最小个体尺寸限制,通常略低于成熟时的平均尺寸。因此,各关键物种之间的捕捞强度并不均衡,且在同一物种内的捕捞也不均衡;相反,存在强烈的“传统”尺寸选择性(古勒斯塔德等人,2014)。
为了研究BH渔业的影响,我们将使用一个针对北欧和巴伦支海(以下简称诺巴模型)进行参数化和调优的端到端阿特兰蒂斯生态系统模型(富尔顿等人,2011),该模型由汉森等人(2016,2019)提供。通过在1980年至2030年期间运行为期50年的模拟情景,我们研究了以相对于生产力的捕捞死亡率收获组分时的交互效应。首先,将选定的物种(包括商业性和非商业性物种)逐一暴露于与生产力成比例的捕捞死亡率下,以探究根据平衡收获方式捕捞单个物种所产生的生态系统效应,并识别出对复合群落具有特殊影响的物种。随后,我们逐步推进到全面实施阶段,设置多个物种同时接受平衡收获的联合运行,以评估BH制度的累积效应。此外,对所有年龄结构群体应用了渔具选择性选项,以在物种内的不同年龄组之间实现平衡。
平衡捕捞此前已在多物种模型中部分研究过(邦迪等,2005;加西亚等,2012;科尔丁等,2016;希思等,2017),但本研究是首次尝试在大西洲模型中基于生产力实施渔业死亡率的BH制度。这也是首个模型应用以研究平衡收获在挪威和巴伦支海的生态系统效应。
材料与方法
亚特兰蒂斯模型
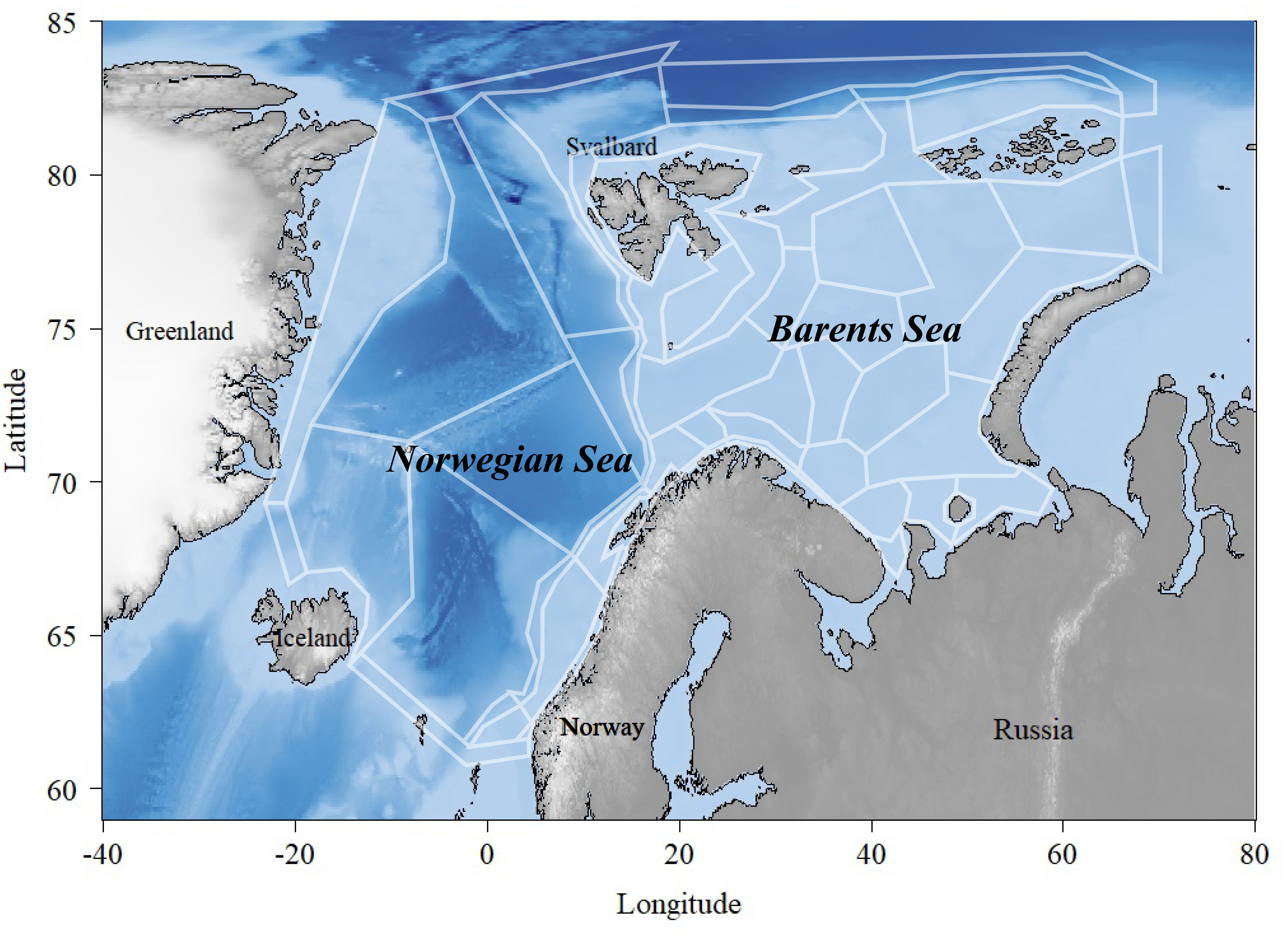
亚特兰蒂斯目前被认为是海洋生态系统中最先进的“如果”情景模型之一(普拉加尼,2007)。该模型模拟了生物地球化学和社会经济过程的空间变化。诺巴模型的区域被划分为60个多边形,覆盖总面积为400万平方公里的北大西洋和巴伦支海,最多包含七个深度层,具体取决于总深度(图1)(汉森等人,2016)。巴伦支海是一个相对较浅的陆架海,平均深度为230米,位于挪威和俄罗斯以北,而挪威海则平均深度大得多,为2000米,位于挪威、冰岛和斯瓦尔巴之间(萨克斯高等人,2009)。挪威海的浮游生物部分生物多样性相对较低,主要由大量洄游鱼类种群主导,例如挪威春季产卵鲱鱼(大西洋鲱)、鲭鱼(大西洋鲭鱼)和蓝鳕(蓝鳕)。另一方面,巴伦支海尽管地处高纬度,却具有相对丰富的生物多样性。这里拥有世界上最大的鳕鱼种群(大西洋鳕鱼),以及其它重要的商业物种,如黑线鳕(黑线鳕)、青鳕(狭鳕)、格陵兰大比目鱼(格陵兰大比目鱼)、毛鳞鱼(毛鳞鱼)、红鱼(Sebastes spp.和挪威红鱼)以及对虾(北方对虾)。
目前,诺巴模型包含53个物种和功能组(表1),它们通过摄食矩阵相互连接。大多数脊椎动物物种具有年龄结构,而无脊椎动物则被归入生物量池中。亚特兰蒂斯不计算多边形之间的水流,而是使用海洋学模型的输出结果。诺巴模型采用自下而上的驱动方式,输入区域海洋建模系统(ROMS:什切佩金和麦克威廉姆斯,2005)提供的温度、盐度和洋流时间序列数据,覆盖东北大西洋(斯科根等人,2007)。收获子模型处理人类对海洋生态系统的开发,重点关注捕捞船队的动态。该模型允许多个船队各自具备特定的特征,如渔具选择性、目标物种和管理体系(富尔顿等人,2011)。诺巴模型包含27种渔业,即所谓的渔业类型(里德等人,2016),每种均有不同的特征和商业捕捞的目标物种(汉森等人,2019)。
实施平衡捕捞
实施BH需要所有物种的生产量或生产力信息,但文献并未提供如何利用这些信息设定捕捞死亡率的明确答案。关于捕捞死亡率应与希思等,2017;周等,2019)存在持续争论,是应与生产力(P/B,单位为“每单位时间”,如本研究中所示)成比例,还是应与生产量(P,单位为“单位时间质量”)成比例。两者的关鍵区别在于,生产量(BH1)是密度依赖的,而生产力(BH2)可以是密度无关的(方程1和2):
Production(BH1):
$$ F(x)= c \cdot P(x)= c \cdot g(x) \cdot B(x) $$
Productivity(BH2):
$$ F(x)= c \cdot \frac{P(x)}{B(x)} = c \cdot g(x) $$
对于这两个方程,物种F的捕捞死亡率x由开发常数c的大小和物种特异性产量P(x)决定,后者由生物量B和生长g计算得出。由于按BH1比例进行捕鱼具有密度依赖性,当生物量较低时捕捞量往往也较低,从而保护物种免于崩溃。相反,根据BH2进行的捕鱼对当前生物量的敏感性较低,因此允许物种被开发至灭绝,正如本次检验的结果所示。
Heath等人(2017)认为,由于平衡收获是一种旨在维持海洋生态系统物种丰富度的生态系统捕鱼方法,因此应在BH1中应用并推荐密度依赖的捕捞死亡率。我们采用了加西亚等人(2012)中描述的方法,其中总产量被定义为个体生长加上补充量,即每年产生的生物量。将总产量除以相应的生物量,得到“人均”生产力速率,通常称为P/B比率。这种将捕捞死亡率设定为与生产力或P/B比率成比例的方法,是平衡收获科学家提出的若干替代方案之一(Jacobsen等人,2014;科尔丁等,2016;周等,2019),也是本研究选择采用的方法。需要注意的是,还存在其他可能的表述方式(有关讨论参见周等,2019)。
与使用P/B比率(或总死亡率)作为输入参数的ECOPATH模型不同,该参数未包含在大西洲模型中。因此,P/B比率的计算是通过利用本研究之前为表征历史渔业而进行的初步运行所生成的生长和生产量输出完成的。大西洲模型有多种应用捕捞死亡率的方式。为此,最佳选择是设定每日收获一定比例生物量的渔业诱导死亡率。为了捕捉生产力的年际变化,每年均计算了P/B比率。从模型输出中提取了有关生长、体重和数量的信息,以估算年龄结构脊椎动物群体的生产力和生物量。对于无脊椎动物,生产量直接取自模型输出。然后,计算所选各组分每年的P/B比率,并通过以下方程转换为比例性捕捞死亡率:
$$ F_{BH}= c \cdot \frac{P}{B} $$
基于平衡收获的捕捞死亡率$F_{BH}$,等于以每年吨数表示的生产力(P)除以以吨数表示的生物量(B),再乘以一个无量纲常数c,该常数决定开发强度。根据卡迪马估计器(特罗阿德克,1977),已提出多个开发强度值,主要范围为0.2至0.4(谢泼德,1982;贝丁顿和库克,1983;保利,1984;加西亚等,1989;斯帕雷和维内马,1998)。然而,决定遵循科尔丁(1993)并采用相对保守的常数0.25,相当于捕捞种群总年产出的25%。然而,在模拟运行过程中显现出,尽管该捕捞率对所有商业物种是可持续的,但对大多数非商业物种而言,25%的捕捞率过高。因此,这些物种的$F_{BH}$被减半至12.5%,以避免立即崩溃(表2)。
对所有年龄结构群体(表2)应用了特定体型选择性,依据为在模拟年份(即1980–2030)期间该年龄组的平均生产力。选择基于体长的逻辑选择性曲线作为选择性选项,因为它能够根据具有年龄结构的物种内部的生产力水平,对不同年龄组施加不同的捕捞压力。选择性曲线通常遵循S形曲线的形状,范围从0到1,其中在不同体长下的保留可能性从0%到100%(斯帕雷和维内马,1998)。该曲线表示为
$$ p_{sel_i}= \frac{1}{1+ \exp(-sel_b \cdot(L - l_{sm}))} $$
物种i的选择性曲线($p_{sel}$)由拐点($l_{sm}$)决定,即在50%选择性时的长度,此时50%逃脱而50%被保留,$sel_b$用于确定曲线的陡度,以及不同年龄组的长度(L),单位为厘米。由于生产力通常随体型大小增加而下降(Peters, 1986),因此预期选择性曲线呈下降趋势,且$sel_b$为负值,以对年轻高产年龄组施加更大的捕捞压力。亚特兰蒂斯使用长度‐重量关系来转换为长度,因为其同生群是基于重量的。为了确定$l_{sm}$和$sel_b$的合适取值,必须通过求解长度‐体重关系方程(Hile, 1936;Martin, 1947)关于长度的表达式,计算各物种在不同年龄的平均长度:
$$ W= a \cdot L^b \rightarrow L= \sqrt[b]{\frac{W}{a}} $$
体长(L)以厘米计,由来自文献的两个物种依赖性参数(a)和(b)确定,并应用于模型中(Hansen et al., 2016),而年龄体重(W)以千克计,则取自模型输出结果。采用非线性最小二乘回归方法来确定$l_{sm}$和$sel_b$的最佳值。通过假设$l_{sm}$和$sel_b$的初始起始值,并应用选择性曲线方程(公式4),选取使选择性曲线最接近生产力水平的$l_{sm}$和$sel_b$值。这与传统的渔具选择性曲线不同,传统曲线旨在保护幼体,即较小的体型大小,并针对较大的体型大小。
被选为进行平衡收获的物种和类群列于表2中。这些物种要么是已被商业开发的物种(物种1–11),要么是非商业物种,基于相对丰富、易于捕捞且是良好的食物来源而被选择(物种12–21)。在对照运行中,非商业组成部分未被收获,包括在挪威渔业中轻微捕捞(例如哲水蚤、小须鲸)或完全未开发的物种(例如中层鱼类、水母)。像浮游植物(难以捕捞)、珊瑚(不可食用)和北极熊(受保护)这样的物种在本研究中被排除在外。
实验设置首先追踪BH对单一物种的单独影响(在补充材料中展示并简要描述如下),然后再逐步进行多物种同时实施BH的全面实施。这样做的目的是研究BH对单个物种的独立影响,以及在BH制度下对多个物种进行捕捞的累积效应。此外,将BH扩展到目前未捕捞(或轻度捕捞)的物种存在明显实际困难,因此我们将其与主要商业物种分开进行考察。三项组合实验为:
(1) “商业物种的BH”,即所有商业物种均采用平衡收获,而非商业物种未被收获;
(2) “非商业物种的BH”,即非商业物种根据平衡收获进行收获,而商业物种则按历史水平收获;
(3) “所有渔业中的BH”,即商业性和非商业性物种均采用平衡收获。
表3给出了本文将用于分析的所有运行的完整列表。所有运行均通过对对照运行进行捕捞努力量调整,并为所有脊椎动物类群添加选择性曲线来完成。未实施平衡收获的商业物种,其收获依据对照运行中的捕捞死亡率(表2中的$F_{Histo}$)进行,且具有应用了平坦的恒定选择性。结果通过生物量(吨湿重)、渔获量(吨湿重)以及一组基于费伊等人(2019)的指标的变化呈现。每公斤渔获价格来自鲱鱼销售联盟(挪威鲱鱼销售联盟,2019)的中上层组分和生鱼协会(挪威生鱼协会,2019)的底层组分。营养级基于科尔等人(2016)所采用的数值。所有绘图均通过“R studio”(RStudio团队,2015)在3.5.2版本下完成。
结果
平衡收获对生物量的影响
挪威海的商业渔业主要由大型中上层鱼类种群主导,包括鲭鱼、蓝鳕和挪威春季产卵鲱鱼(下文简称鲱鱼)。在巴伦支海,主要的商业物种为东北北极鳕鱼(下文简称鳕鱼)、毛鳞鱼、黑线鳕、青鳕、格陵兰大比目鱼、喙红鱼、金红鱼、红鱼和对虾。所有商业物种的捕捞死亡率与生产量之比($F_{BH}$)的平均值如表4所示,同时列出的还有对照运行中的历史捕捞水平平均值($F_{Histo}$)以及基于最大可持续产量的捕捞水平($F_{MSY}$)。
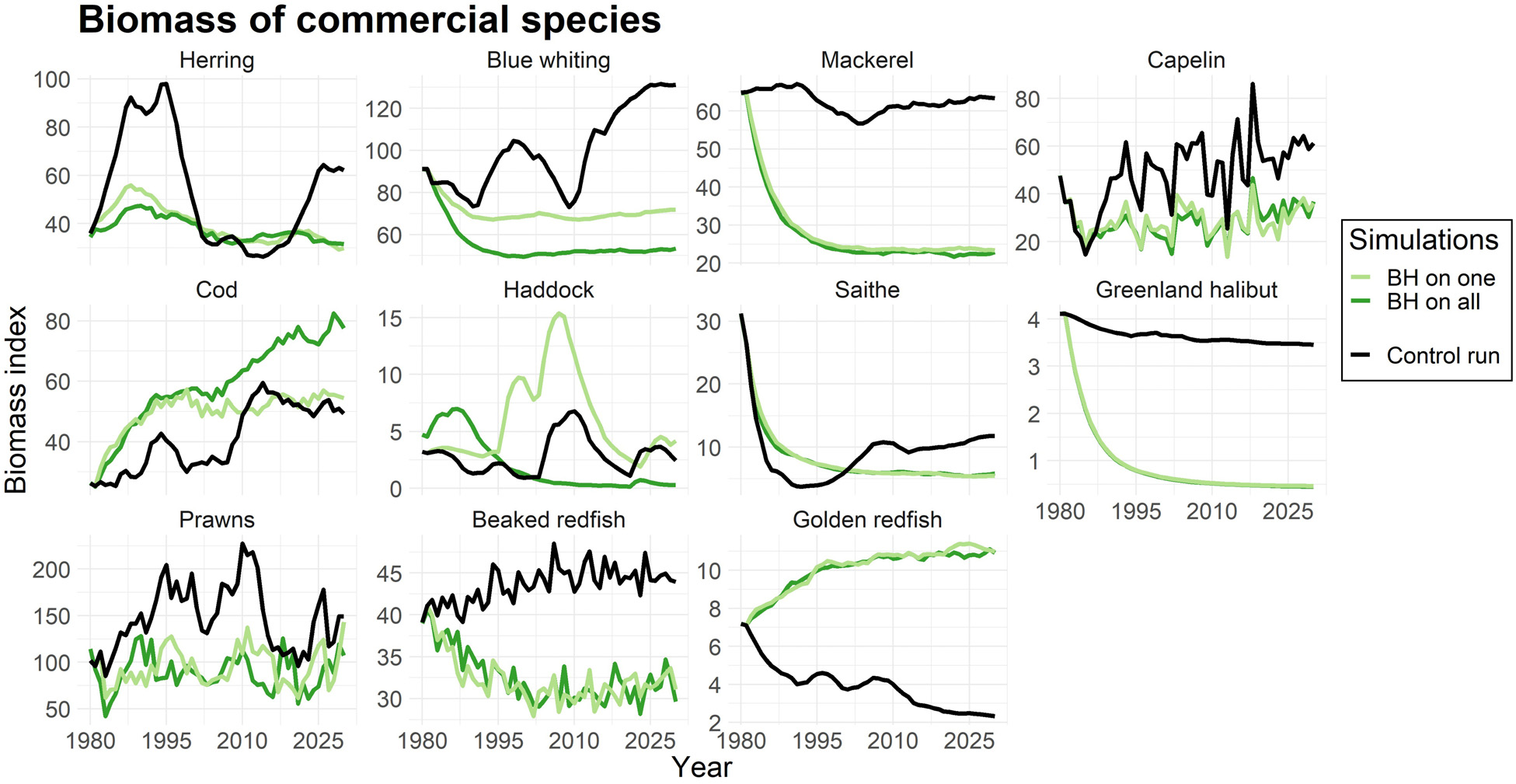
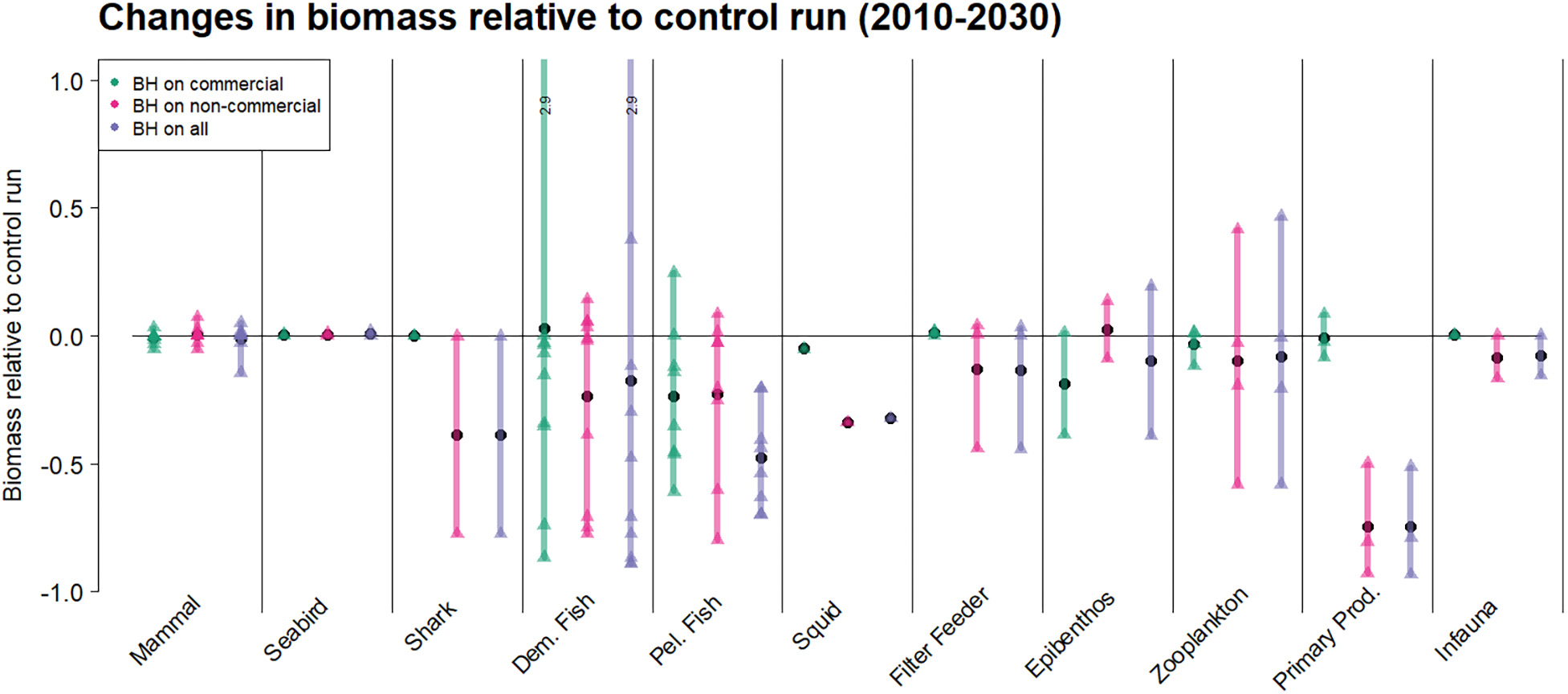
为了评估BH制度对每个物种的直接和间接影响,我们进行了模拟实验,其中仅有一种物种受到平衡收获的影响,随后所有选定物种均在$F_{BH}$水平下进行捕捞。图2展示了商业物种的生物量:(i)仅一种物种受到平衡收获时(浅绿色线);(ii)所有选定物种均受到平衡收获时(深绿色线)。黑线表示应用传统渔业方式的对照运行中的生物量。
正如预期,图2反映了捕捞死亡率变化对生物量的影响(表4)。捕捞死亡率较低的物种(如鳕鱼和金红鱼)生物量有所增加,而由于捕捞压力大幅上升,鲭鱼、格陵兰大比目鱼和对虾的生物量则显著减少。
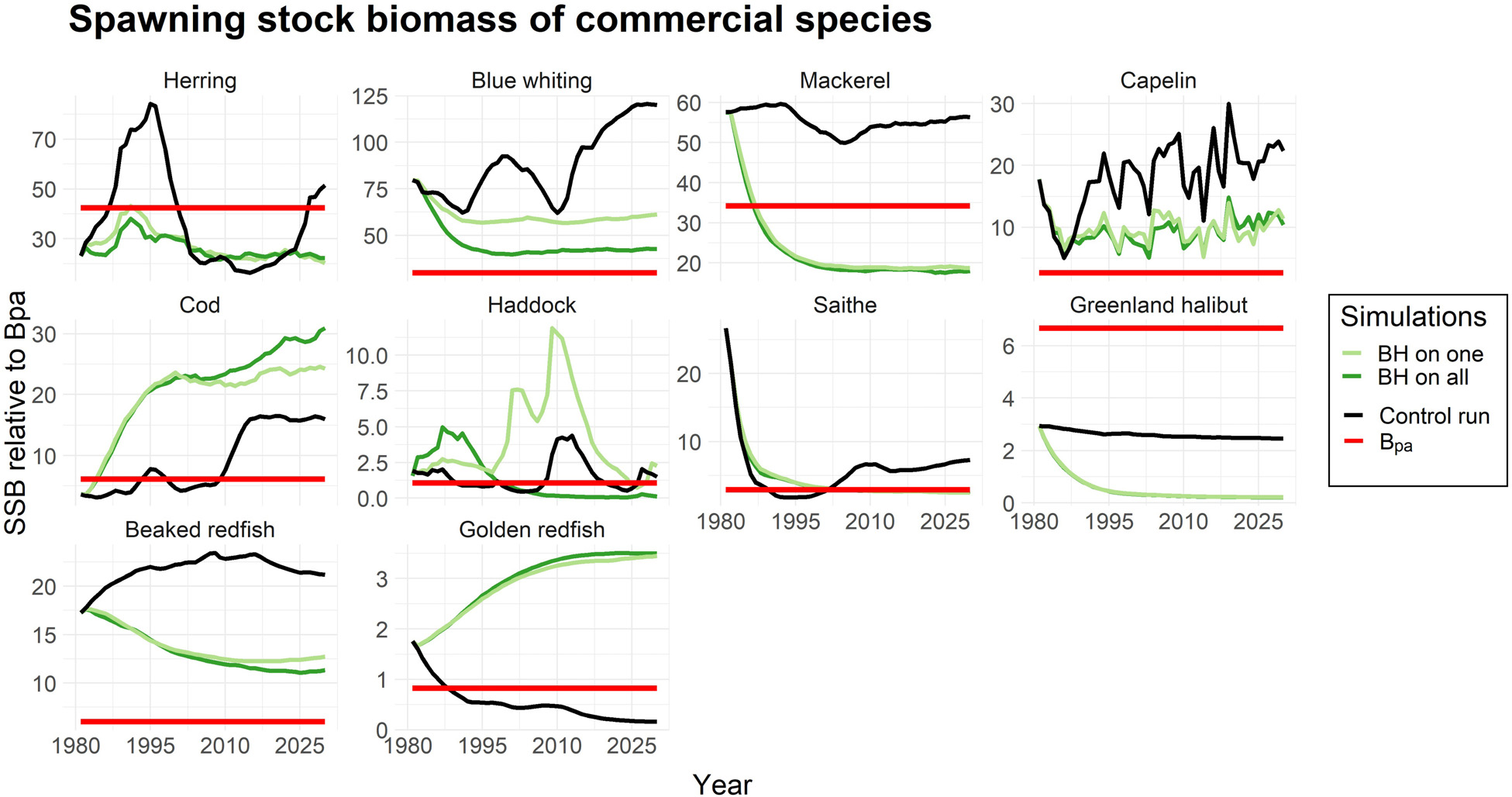
然而,由于捕捞压力增加而导致的生物量下降,只要不导致补充过度捕捞,对资源群体而言并不一定是严重的。为了评估这一点,我们绘制了各物种的产卵群体生物量(SSB)以及bpa,即预防性参考点(ICES, 2017, 2018a),当资源低于该点时将面临补充过度捕捞的风险。图3显示,尽管在BH情景下蓝鳕、毛鳞鱼和喙红鱼的生物量大幅减少,但其SSB值仍高于bpa。相比之下,在BH制度下,鲭鱼和鲱鱼的SSB值被降至bpa以下。金红鱼则是一个例子,在对照运行中的传统捕捞制度已导致其SSB处于极低水平。
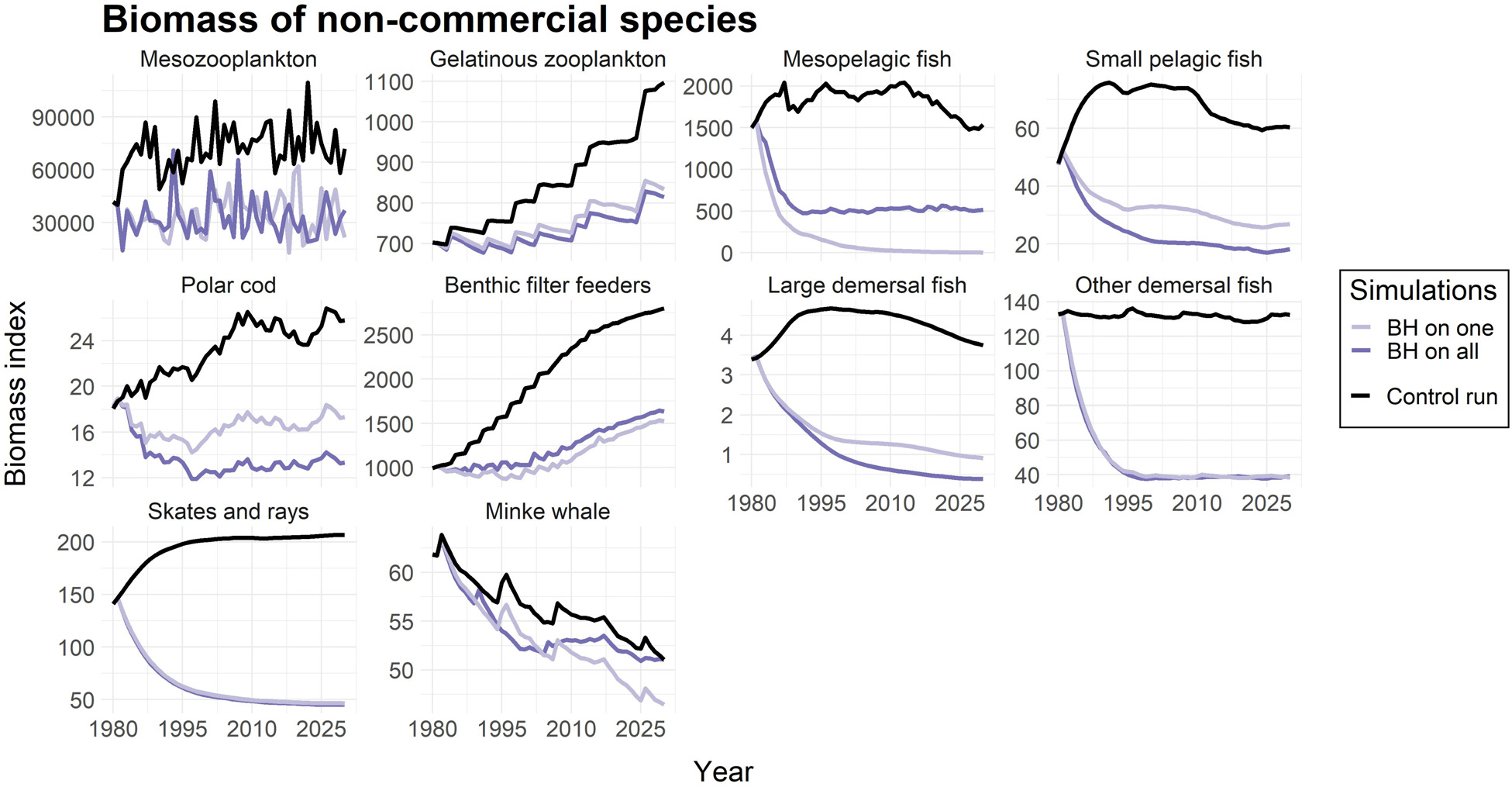
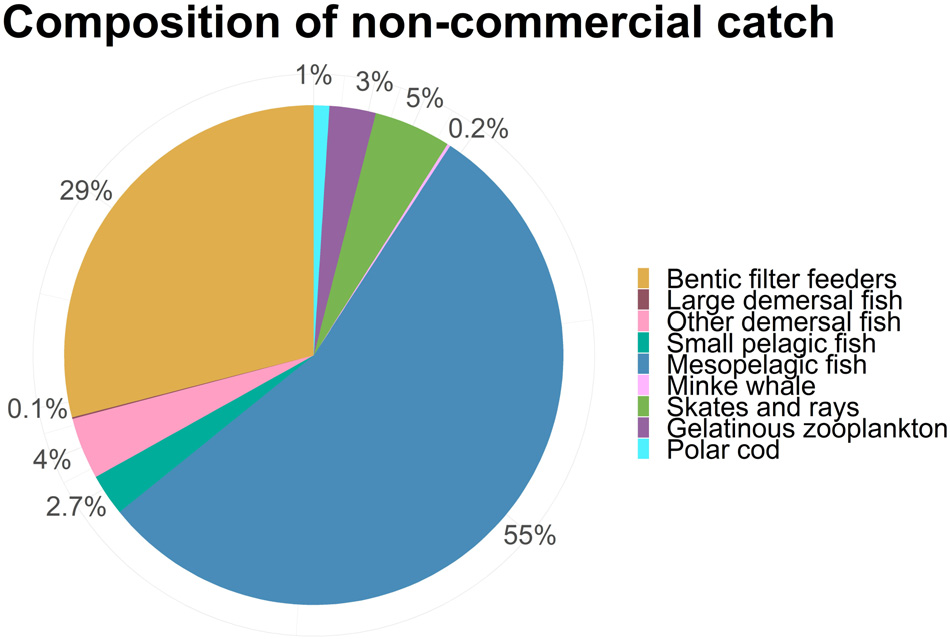
该研究还包括了对被视为“非商业性”且在对照运行中未被针对的物种进行捕捞。这些物种的计算捕捞死亡率($F_{BH}$)列于表4中。图4显示,所有非商业种群在受到平衡收获制度影响时生物量均有所下降,这与受到捕鱼影响时的预期一致。然而,不同物种的减少幅度差异很大。中层鱼类几乎濒临崩溃,两类底栖鱼类(包括蓝ling、牙鳕和狼鱼)以及鳐鱼和魟鱼的生物量减少了近75%。中型浮游动物和小型中上层鱼类的减少程度较小,约为50%,而胶质浮游动物、极地鳕、底栖滤食者和小须鲸受影响更小。
利用亚特兰蒂斯生态系统模型探索北欧和巴伦支海的平衡捕捞
结果
平衡收获对渔获量的影响
平衡捕捞旨在提供更高的产量,同时比传统的选择性捕鱼更好地保护生态系统结构与功能。在分析的第二部分,我们探讨了不同BH情景下对渔获量的影响。估计的年总渔获量以模拟最后20年的平均值表示(2010–2030年)。
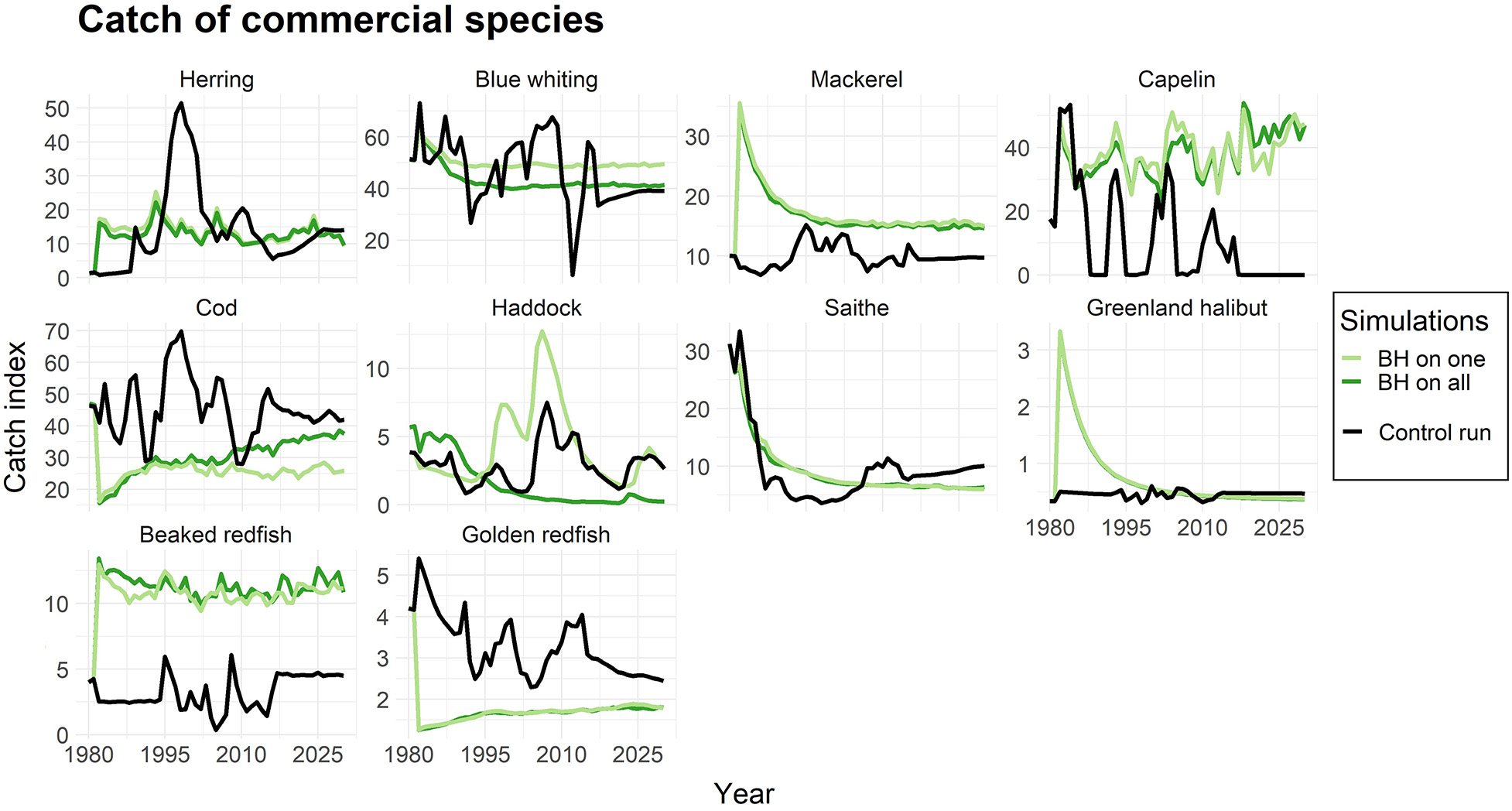
图5展示了商业物种在单独实施平衡收获以及所有物种共同实施平衡收获时的渔获量,与对照组相比的情况。结果显示,某些物种的渔获量较高(例如鲭鱼、毛鳞鱼、喙红鱼),而其他物种的渔获量较低(例如鳕鱼、金红鱼)。尽管渔获量在实施BH制度的最初几年内不稳定,但大多数商业物种的渔获量在BH制度下似乎变得更加稳定,进入模拟的情况。
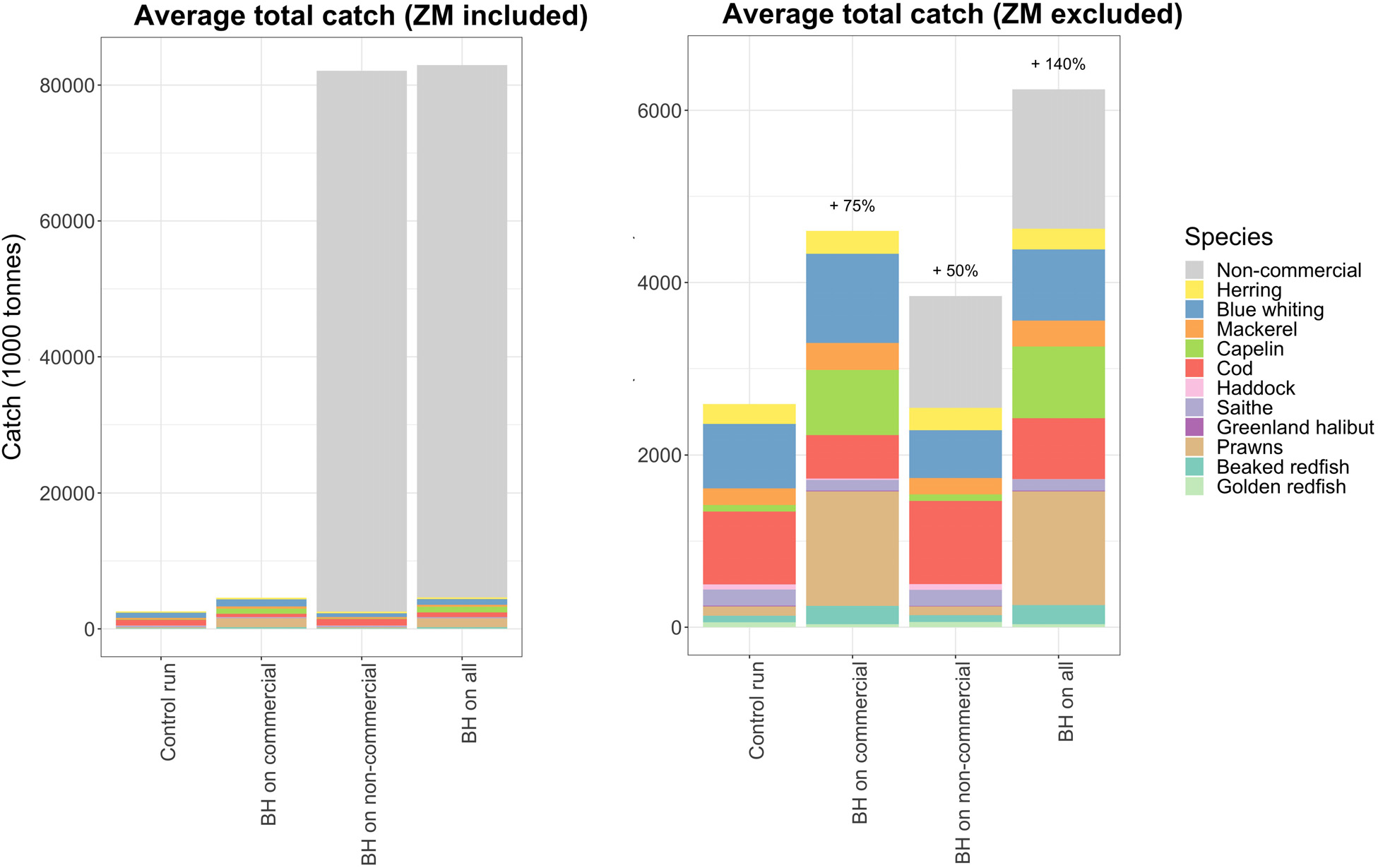
下一个问题是,哪种BH组合(即“商业渔业中的BH”、“非商业物种的BH”或“所有渔业中的BH”)能够产生最高的总产量。将三次组合运行与对照运行进行比较,其中商业物种以历史水平收获,而非商业物种未被收获。采用模拟的最后20年(2010–2030年)的平均总渔获量(此时渔获量已趋于稳定)来评估长期产量。
 包含和(B)不包含中型浮游动物(ZM)。本图中所有非商业物种被归为一类,称为“非商业种群”。)
包含和(B)不包含中型浮游动物(ZM)。本图中所有非商业物种被归为一类,称为“非商业种群”。)
图6显示了在不同平衡收获变化下的总渔获组成。当更多物种被纳入并以BH制度进行收获时,总渔获量增加,但这增加主要由毛鳞鱼、对虾和非商业物种的渔获量上升所致。图6A中将非商业物种的渔获量合并显示,显示出额外近80百万吨的渔获产量。然而,这部分渔获量几乎全部(98%)为中型浮游动物,在图6B中已被排除,以便更方便地比较其余物种。
当仅将非商业物种暴露于BH制度下时,商业物种的总渔获量减少了24,000吨(图6),与新增的80百万吨(主要为低营养级物种)相比,这一减少幅度相对较小。图7显示,商业物种渔获量的减少主要是由于蓝鳕减少了20万吨,但这减少部分被鲱鱼和鳕鱼渔获量的增加所弥补。
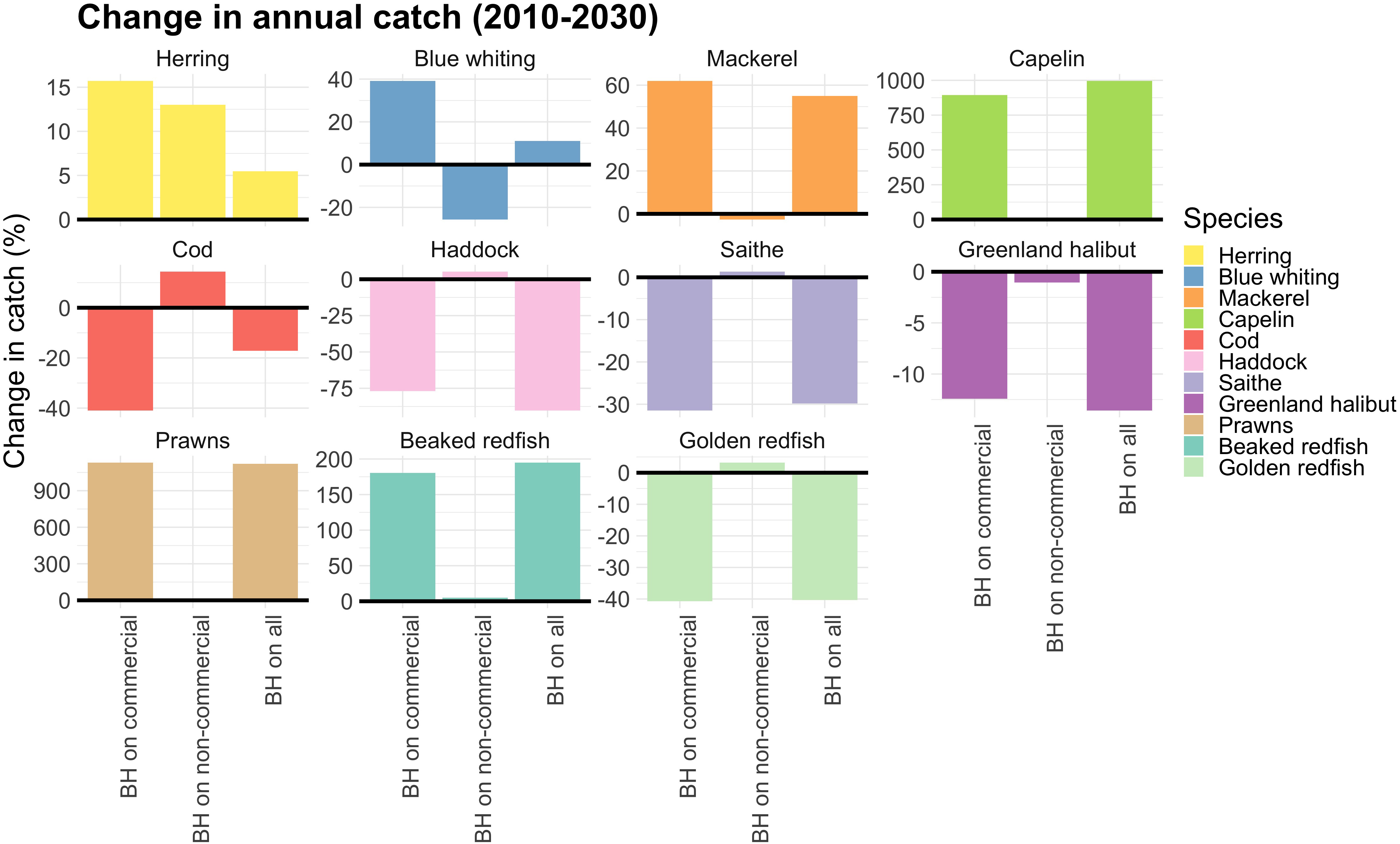
排除中型浮游动物后,非商业物种的新增渔获量约为160万吨。这些新增渔获量主要由中层鱼类和底栖滤食者构成,此外还包括少量底栖鱼类、水母、鳐鱼和小型中上层鱼类(Figure 8)。
平衡收获对生态系统结构的影响
图9展示了每个功能群的生物量变化如何反映整个生态系统的响应。每个功能群所包含的各个物种和功能组可在表1中找到,并在图中以三角形表示。该图显示,包含受平衡收获影响物种的功能群响应最为强烈。然而,当非商业物种受到平衡收获影响时,初级生产者群体似乎受到的影响最大。
为了更好地理解实施BH制度的全部影响,我们需要纳入比渔获量和生物量更多的方面。图10比较了五个额外的指标:(i)渔获物平均营养级(MTLCatch);(ii)生物量平均营养级(MTLBiom);(iii)浮游动物与远洋鱼类的关系(ZooPel);(iv)远洋鱼类与底栖鱼类的关系(DemPel);以及(v)仅商业渔获量的价值,这些指标均基于三种综合情景以及对照运行的结果。“所有渔业中的BH”这一完全平衡运行情景在除渔获物平均营养级外的所有指标上均取得了最高值。相反,历史对照运行则给出了相反的结果,在除MTLCatch外的所有指标上均为最低值。
在商业物种或非商业物种进行平衡收获的情景中,结果介于另外两种情景之间:“商业渔业中的BH”产生了更高的渔获量,而“非商业物种的BH”则导致系统中生物量平均营养级更高。“所有渔业中的BH”情景在浮游动物与远洋鱼类的比率以及底栖鱼类与远洋鱼类的比率方面也得分最高。
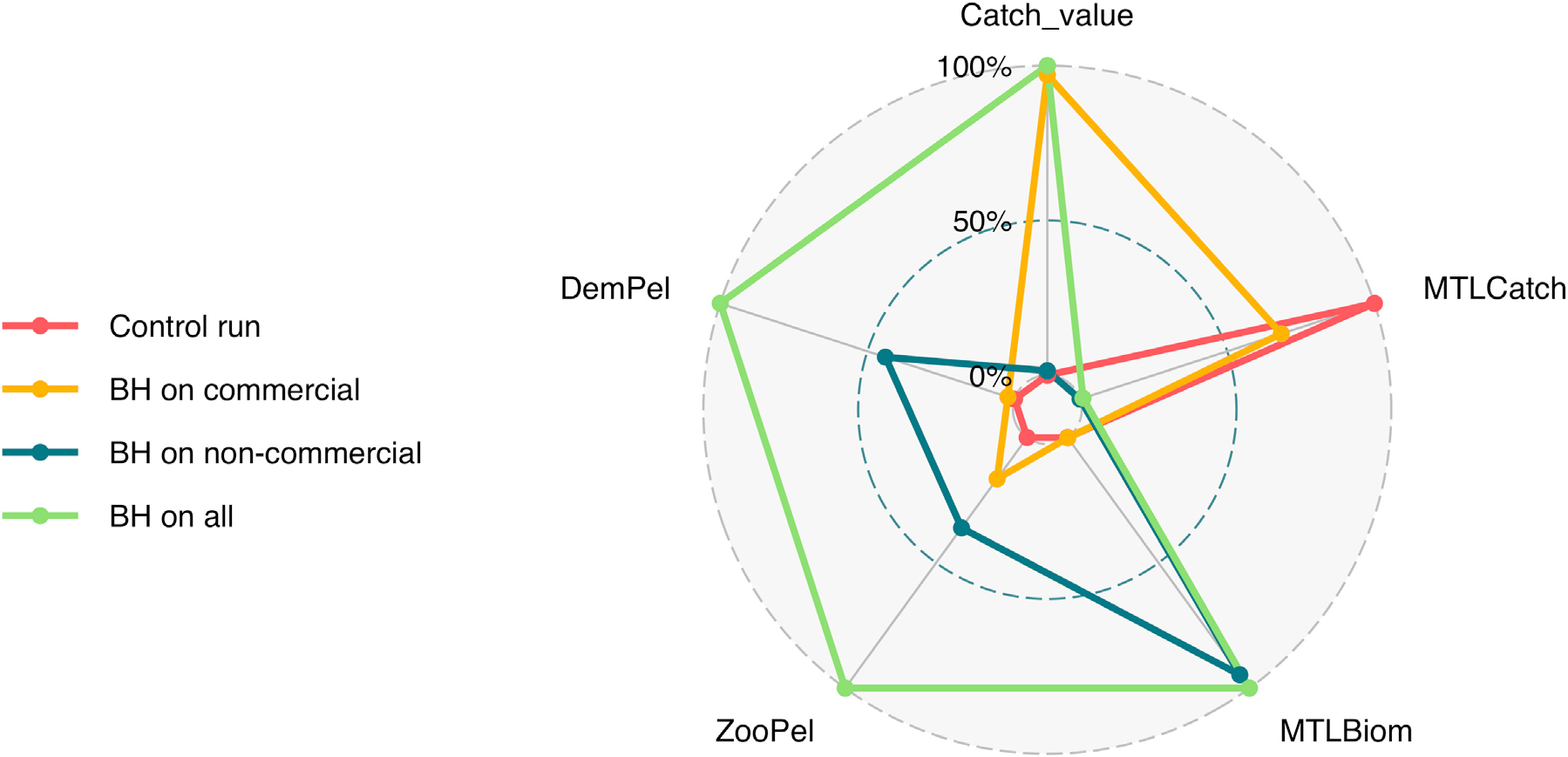 渔获物平均营养级(MTLCatch);(ii)生物量平均营养级(MTLBiom);(iii)浮游动物与中上层鱼类的关系( ZooPel);(iv)中上层鱼类与底栖鱼类的关系(DemPel);以及(v)三种组合情景和对照运行下的仅商业渔获量价值。)
渔获物平均营养级(MTLCatch);(ii)生物量平均营养级(MTLBiom);(iii)浮游动物与中上层鱼类的关系( ZooPel);(iv)中上层鱼类与底栖鱼类的关系(DemPel);以及(v)三种组合情景和对照运行下的仅商业渔获量价值。)
讨论
平衡收获对物种的影响
在对生态系统中的渔业进行广泛调整时,很难区分直接影响和间接影响。根据生产力设定平衡收获(BH)实际上提出了针对每个物种的选择性以及物种间捕捞压力相对关系的改变。鉴于这些变化可以分别实施,因此有必要研究任何变化中有多少来自于对特定物种渔业的调整,有多少来自于改变物种间捕捞压力平衡所带来的间接生态系统效应。
平衡收获对单个物种的影响结果见于补充材料,此处重点介绍一些关键发现。
几乎所有商业物种目前的捕捞水平都接近其各自估计的最大可持续产量,但格陵兰大比目鱼、金红鱼和毛鳞鱼除外(ICES, 2018b,c)。对于格陵兰大比目鱼,当前评估模型仅根据长度数据进行调整,因此对FMSY的估计存在不确定性(ICES, 2015b),这使得直接比较变得困难。平衡收获情景预测的长期渔获量略低于历史捕捞情景,且资源生物量显著更低。目前金红鱼种群处于过度捕捞状态(ICES, 2018b),而平衡收获下的捕捞压力远低于历史捕捞水平。应用这种较低的捕捞压力可实现种群恢复,并最终带来更高的渔获量。
对于鳕鱼,单独对鳕鱼实施平衡收获策略时,长期渔获量低于历史捕捞情景;然而,当将平衡收获应用于整个生态系统的所有组分时,鳕鱼的长期渔获量仅略低于历史捕捞水平。
对毛鳞鱼的模拟平衡收获表明,更高的捕捞死亡率(表4)可带来高达3百万吨的额外产量。然而,在亚特兰蒂斯模型中,毛鳞鱼渔业被设定为恒定的捕捞死亡率,而这种策略对于像毛鳞鱼这类生命周期短且生物量波动大的种群而言效果较差。这与该种群的实际管理方式不符,因为毛鳞鱼的捕捞控制规则是一种所谓的逃逸策略,即保证一定数量的个体能够成功繁殖,仅允许捕捞超出部分,从而允许年际间产量出现较大波动。这一动态当前阿特兰蒂斯模型对渔业制度的模拟不够准确,因此该种群与实际渔业的比较存在问题。
这四个种群凸显了本研究中所进行建模工作的若干注意事项,以及考虑各个物种动态的重要性。首先,当种群状态和参考点不明确时,建模和比较结果将变得不确定。其次,对于当前处于过度捕捞状态的种群(如金红鱼),任何降低捕捞压力的措施都可能带来益处。在这种情况下,平衡收获(BH)有助于种群恢复,但同样可以通过不采用BH的方式来实现捕捞压力的降低。总体而言,这表明在模拟测试BH相对于良好实践的传统管理的优势时,应将其与管理良好的渔业进行比较,而不是与当前资源枯竭的种群状态进行比较。对于鳕鱼这类捕食者,比较仅改变鳕鱼的捕捞与改变整个生态系统的捕捞之间的差异,突显了生态系统层面效应可能部分抵消(或潜在加剧)预期渔获量损失的可能性。最后,毛鳞鱼是一个渔业管理在大西洲模型中未能很好体现的实例,因此其结果应谨慎对待。此外,毛鳞鱼也是高度变动的短生命周期物种的一个例子,在这种情况下固定捕捞压力是一种不佳的捕捞策略,因此也是需要慎重考虑采用BH策略的典型案例。
更多单个物种的详细信息见Supplementary Material,但本概述旨在强调,单个物种动态对于将平衡收获应用于整个生态系统的结果至关重要,因此必须使用能够解析此类细节的模型工具。
平衡收获对总渔获量的影响
在将对照运行与多种物种的综合平衡捕捞制度结合运行进行比较时,结果表明,受到平衡捕捞影响的物种越多,总体渔获量就越高。然而,商业物种中的主要增长来自毛鳞鱼和对虾,这两种物种应当谨慎对待。在大西洲模型中模拟对虾似乎是一个众所周知的问题(B. 弗ulton,个人交流),即使在种群评估中,生物量估计也存在很大的不确定性(ICES, 2013)。如前所述,在诺巴模型中模拟毛鳞鱼渔业也存在困难,该模型无法容纳逃逸规则策略。毛鳞鱼还具有复杂的生活史策略,其产卵后的死亡率非常高,因此在解释结果时需要格外小心。
当将中型浮游动物纳入综合平衡捕捞制度时,它几乎完全主导了潜在的总渔获量(图6A),每年接近8000万吨,是当前挪威总渔获量的20倍(渔业管理局,2016年)。对中型浮游动物的捕捞压力设定为生产力的25%,导致其生物量减少了50%(图4),但如此大量地捕捞中型浮游动物对其他物种的影响却出乎意料地小。现实中近8000万吨的捕捞量并不可行,但考虑到当前配额仅为16.5万吨(渔业管理局,2016年),而该资源的现存量生物量为3000万吨,年产量估计为2.9亿吨,因此增加配额可能不会产生明显的影响。本研究直接针对每个建模物种设定捕捞压力,并未考虑因渔获量变化而导致的其他物种的兼捕情况。因此,我们未考虑如此大规模增加中型浮游动物捕捞可能导致的对其他物种的卵和幼体兼捕增加所带来的潜在影响。
另一个有趣的结果是,在BH制度下,大多数商业物种的年际渔获量波动小于传统捕捞制度,表明平衡收获将产生更稳定的渔获产量。这反映了捕捞死亡率的变化,因为在对照实验中,$F_{BH}$比$F_{Histo}$更稳定。
模拟结果表明,根据当前模型,在管理良好的巴伦支海和挪威海,平衡收获对已进行商业开发的种群带来的收益相当有限。尽管BH制度所建议的更高捕捞强度使许多物种的渔获量增加,但这以显著降低现存量生物量(图2)和后续资源为代价,单位捕捞努力量渔获量下降,以及补充过度捕捞风险增加(4),少数例外情况(例如,喙红鱼)。
然而,这并不一定意味着平衡收获是一种错误的想法。豪厄尔等人(2016)基于斯卡雷特和皮彻(2016)的Ecopath模型,研究了挪威海和巴伦支海中28种捕捞物种的产量与生产量之间的关系。他们得出结论:挪威和巴伦支海目前的捕捞管理制度已经相当平衡,且比大多数其他海洋系统更为平衡(科尔丁等,2016)。这支持了以下发现:任何额外产量预计将主要来自目前未开发或开发不足的物种。
平衡收获对生态系统结构和渔获量价值的影响
在考虑渔获量总价值时,“所有渔业中的BH”和“商业渔业中的BH”情景产生的价值最高。这主要来自对虾,其价值和渔获量均较高,此外还包括毛鳞鱼和喙红鱼。喙红鱼是一种相对高价值的物种,因此其渔获量几乎大幅增加。毛鳞鱼的价值相对于其他物种较低,但在平衡收获(BH)情景下,其渔获量比其他情景高出10倍以上,从而提升了总价值。然而,如前所述,对虾和毛鳞鱼的结果应谨慎对待。此外,还应考虑到非商业物种捕捞可能带来的经济效益,尽管本文未进行此类估算,因为相关价格和市场信息尚不明确。
在所有平衡捕捞制度下,渔获量的营养级均为最低,这是符合预期的,因为平衡收获(BH)的核心理念之一就是增加对较低营养级物种的捕捞。然而,这可能会带来问题,因为当前市场通常更青睐较高营养级的物种。根据各物种生产力比例施加的特定体型选择性,通常意味着与传统渔业制度相比,对幼鱼的捕捞压力更大。显然,在诺巴区域捕捞较小个体的鱼类在当前市场上售价也会更低,从而带来经济成本(本研究未考虑这一点)。从生态角度来看,对幼鱼增加的捕捞死亡率(F)实际上是对选择性曲线的一种调整,使其比目前的传统渔具选择性曲线更贴近自然死亡率(和生产力),因此我们预计这将减少渔业诱导进化方面的潜在影响(劳和普兰克,2018年)。由于平衡捕捞制度包含了对非商业物种的收获,导致低营养级物种(如中型浮游动物)从系统中被更多地移除,因此总生物量的平均营养级相应显著提高。
选择ZooPel和DemPel的比率是为了研究生态系统结构在BH情景下可能发生的变化。“所有渔业中的BH”情景中浮游动物与中上层鱼类的相对数量最大。这一点有点令人惊讶,因为该情景包含了对中型浮游动物的巨大收获。然而,从图9可以看出,中型浮游动物的移除部分被其他浮游动物物种(主要是小型浮游动物)的增加所补偿,这可能使整个浮游动物生物量保持稳定,而中上层鱼类则被更强烈地捕捞。底层鱼类与中上层鱼类比率在全面实施BH的情景中也是最高的,这里也主要是由于中上层鱼类的减少,而非底栖鱼类的增加所致。对于这两个指标而言,“所有渔业中的BH”情景下的比率相比对照运行翻了一番,表明中上层鱼类的数量相对于浮游动物和底栖鱼类减少了一半。
移除如此大比例的“中间”营养级可能会随时间改变生态系统的结构,并带来未知后果,应当予以考虑。因此,结果表明,在挪威和巴伦支海实施全面的BH制度,以总产量25%的开发水平进行开发,可能导致已被开发物种的生物量进一步下降,其所带来的总产量收益可能无法弥补这种影响。另一方面,扩大渔业目标至目前尚未商业开发的物种,特别是较低营养级的物种,可能会提供可观的额外产量,尤其是来自中型浮游动物的产量。然而,较低营养级的物种通常经济价值较低,大量移除特定营养级可能给生态系统带来未知的结构性变化,这一点需要加以考虑。
不确定性与未来研究
必须根据所用模型的假设和局限性来评估结果。尽管大西洲模型能够捕捉生态系统中广泛存在的变异性,但这种复杂性的增加也随之带来了更大的不确定性(豪厄尔等人,2016)。为了弥补对过程认知的不足以及相关数据的缺失,不得不做出若干假设和“估算”(Fulton,2010;汉森等人,2016)。普遍性、精确性和真实性是模型中三个理想特征,但不幸的是,高度复杂的模型通常具有大量参数和高分辨率,一般难以同时实现这三个特性,并往往倾向于弱化其中一项以优化另外两项(Olsen等,2016)。作为一种端到端模型,大西洲模型旨在提供一个整体背景,但本研究中显然发现了一些弱点和难以解释之处。
一个不足之处在于,在模型的参数化和调优过程中,非商业功能组存在更大的不确定性,这既是因为关注重点通常放在“重要”的商业物种上,也是因为关于非商业组的信息较少(塞西莉·汉森,个人交流)。改进这些结果的第一步是对非商业物种进行全面重新调优,以实现所有功能组之间恒定的BH捕捞死亡率,而无需像本研究中那样进行临时性调整(表2)。所选择的开发水平为估计生产力的25%,这被认为是比较谨慎和保守的,因为即使对于饵料物种,通常认为高达40%的开发水平仍是可持续的(Patterson,1992;Pikitch等,2012)。因此,在当前模型中,为了避免大多数非商业物种的崩溃而需要将其开发水平降低至12.5%,这表明这些物种的参数化和模型调优可能不如商业物种准确和稳健。
应重新审查无脊椎动物的生产量和生产力水平的计算方法,因为这导致对虾和中型浮游动物的$F_{BH}$值非常高。无脊椎动物的生产量计算方法与脊椎动物不同,无脊椎动物被归入具有特定生产力的生物量池,并通过总面积进行外推。
除了将本研究扩展到其他生态系统外,还应将其扩展到包括各种类型的平衡收获(BH)。如前所述,目前提出了两种类型的BH(公式1和2),在类似亚特兰蒂斯的模型中比较基于生产量的平衡收获(BH1)与基于生产力的平衡收获(BH2)将十分有意义,以验证假设的BH1对防止物种灭绝的保护作用是否成立。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









