







序列空间充满了可折叠成紧凑蛋白质结构的序列
现在,让我们来探索随机序列的本质。任意的氨基酸序列会折叠成类似天然的结构吗?这个想象的序列空间汤(Sequence-space soup)具有包含所有序列空间的统一表示。这是一种非常好的概念可以用于解决随机氨基酸序列结构与性质之间的关系。
你可能会人为这样的一个序列空间汤可能是一个特别荒凉无趣的空间。它里边的链可能都是打开的,非折叠的或者不具有结构特征-与折叠的蛋白质结构没有任何相似之处。相反,HP的建模却展示出序列空间汤的性质是相对类蛋白化的(BOX 7.3)。在这种空间的分子往往是非常致密的且具有二级结构。这个现象来自于一个现象:(1)超过一半的天然氨基酸都是具有疏水性的,因此任何给定的序列都有比较可观的疏水氨基酸残基,(2)因此这些链在水中的时候往往会形成一个疏水性的中心,(3)由此产生的致密性可以有助于稳定那些氢键相连的特定二级结构并且同时可以确保那些致密链的结构。
====================================================
BOX 7.3 随机序列具有类蛋白化的概率有多少?
进入随机序列并拉出一个折叠成蛋白质结构的序列的概率是多少?考虑所有具有N = 100残基的蛋白质的序列空间汤。序列的数量是20100个。我们需要考虑两种不同的计算方法。首先,找到一个特定序列的概率是
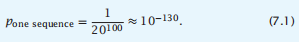
第二,找到任何具有HP模式并折叠成特定结构的序列的概率是:
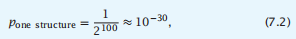
我们取2而不是20,因为折叠代码主要是H和P。此,通过随机搜索,找到一个具有正确的疏水和极性氨基酸模式的序列,需要指定一个特定的天然折叠,比找到一个特定序列的可能性高100个数量级(图7.13)。另外,我们甚至可能不需要在蛋白质表面使用完全正确的HP模式,因为蛋白质表面通常对突变更容易耐受。这些H,P代码参数与涉及阻遏物核心突变的实验以及序列随机化实验的实验定性一致。这些论点说明了为什么自然蛋白质不是不太可能从随机的氨基酸序列中产生,例如在早期进化中。
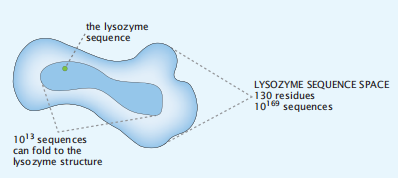
图7.13 在所有可能序列的汤中发现溶菌酶的概率是多少?溶菌酶为N = 130。因此,在所有可能的20130 = 10169序列的汤中,找到溶菌酶的一个序列的概率基本上是零。但是,找到任何具有正确的疏水-极性模式的序列来折叠成溶菌酶的结构的可能性都要高得多。它在(1/2)(130/3)=10−13附近,说明在核心中大约有N/3个残基,它应该是疏水的。
====================================================
蛋白质和其他聚合物可以被设计成稳定折叠的结构
近年来,将工程蛋白质和其他聚合物折叠成所需的结构已经成为可能。利用(a)蛋白由氢键二级结构构建的原理,(b)蛋白质的核心大多是疏水的,大多排列良好,而(c)稳定的蛋白质结构很少有埋藏电荷,结合蛋白质数据库中许多天然结构的见解,几个研究小组已经创造了人类设计的蛋白质。图7.14显示了Baker设计和研究的蛋白质Top7。Top7在两个方面是关键的关键进展:(a)折叠成目标结构,(b)这种结构以前在自然界中从未见过。蛋白质也被设计成可以催化反应。
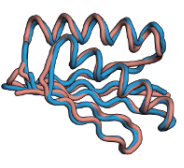
图7.14 蛋白质可以被设计成折叠成预先指定的天然构象。Baker和他的同事们设计了一个氨基酸序列来折叠成蓝色的新模型构象。然后,他们用分子生物学技术生产出这种蛋白质,将其结晶,并确定其结构(橙色)。因此,所设计的序列被证明可以折叠成目标结构。
蛋白质并不是唯一一种可以折叠成特定结构的聚合物。一些RNA分子可以折叠成紧凑而独特的结构。可以折叠成特定稳定结构的聚合物被称为折叠体。被称为肽的聚合物,具有非生物取代甘氨酸主链,被设计成折叠成螺旋束。
-进化论并不是一种抽象的概念,它现在正在真实发生
进化在我们周围进行。它发生在一个细胞或生物体的生命周期中。进化并不一定是缓慢的。癌变肿瘤是体细胞进化的一个例子,即在机体生命周期内发生的细胞特性的变化。寄生虫、传染性病毒、良性和致病菌都在人和动物体内进化和适应。在定向进化中,突变和自然选择的实验室周期被用来在数月到数年的时间尺度上创造具有商业重要性的新蛋白质。
耐药性是在作用中进化的一个例子
耐药病原微生物是一个主要的生物医学问题。一些耐药细菌,如MRSA(耐甲霉金黄色葡萄球菌),有时被称为超级细菌,同时对许多不同的抗菌药物产生耐药性,不受我们现代主要药物的影响。这是产生耐药性的一种方式。首先,考虑一种治疗疾病的药物。这种药物与致病性生物体的关键蛋白质结合,如细菌,杀死该生物体。问题是,这种药物可能会杀死其中一些致病细胞,但可能不会杀死其他细胞,这仅仅是由于种群的自然多样性。种群在药物的存在下进行复制时,耐药细胞现在可以繁殖了。通过这种方式,耐药生物体开始在许多复制周期中主导种群。有些进化的速度相对较快。艾滋病毒和细菌性病原体在不到五年的时间内迅速演变为,对药物产生耐药性。相比之下,一家制药公司首先要发现和开发一种新药,通常需要10-20年的时间,耐药性问题具有严重性。
蛋白质是如何进化出对药物的耐药性的?想想一个致病性生物体;称之为wt(野生型)。想想那个致病性生物体中的一种蛋白质,它具有与天然配体A'结合的正常功能;见图7.15的最上面一行。想想一种药物与相同蛋白质的结合,阻断配体A'的自然作用;参见图7.15的中间一行。上面和中间的几行总结了病原体的正常作用以及药物如何抑制病原体。随着时间的推移,致病生物体可以通过进化蛋白质的突变而对药物产生耐药性。图7.15的底部一行显示了蛋白质的突变,破坏了药物结合,恢复了致病蛋白的正常生物作用。这表明了生物体进化成耐药的一种方式,还有其他方式产生耐药性。
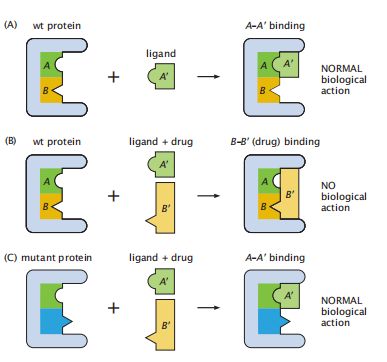
图7.15 致病性蛋白如何产生耐药性?这里有一种机制。(A)病原体的正常功能:天然配体(A)与病原体的野生型蛋白结合,导致其正常的生物学作用。(B) 药物 (B)杀死了病原体:一种药物分子成功地战胜了天然配体,阻止了配体对致病蛋白的作用,杀死了致病生物体。(C)致病蛋白发生突变,抵抗药物的作用:致病蛋白发生突变,改变其结合位点,阻止药物结合,并重新允许致病蛋白的正常作用。
分子时钟:一些进化变化以恒定的速度进行
不同类型的蛋白质往往以不同的速度进化。然而,对于一个给定的蛋白质和该蛋白质中的一个给定的氨基酸位置,突变率通常随着时间的推移而近似恒定,甚至在不同的生物体上也是如此。1962年, Zuckerkandl 和 Pauling将这种突变率的稳定性称为分子时钟假说(molecular clock hypothesis)。例如,在球蛋白中,突变和自然选择导致每位点每十亿年大约一个氨基酸的变化。在进化过程中,一种蛋白质以恒定的速率与另一种蛋白质分离。图7.16显示了分子时钟假说的证据。图7.16的x轴给出了从今天到两个物种分化的时间,有时这是由古生物学确定的。y轴给出了两个序列之间的 Hamming距离,即它们之间的氨基酸差异的数量。从图中可以看出,自蛋白质进化分化以来,不同蛋白质中氨基酸变化的数量近似为时间的线性函数。
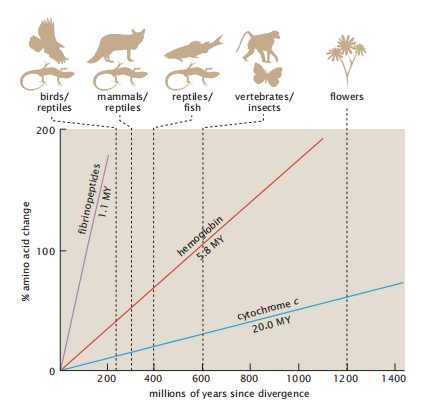
图7.16 蛋白质以恒定的速率变化,如“进化时钟”。对于每一种给定类型的蛋白质,氨基酸变化的数量以恒定的速率线性增长。对于不同的生物体,这个速率是相同的。时钟速率取决于蛋白质的类型。例如,纤维蛋白肽比血红蛋白进化得快,而血红蛋白比细胞色素c进化快。
图7.16还显示了不同蛋白质的分子时钟以不同的速度跳动。在一个给定的蛋白质中,不同的氨基酸位置可以以不同于其他氨基酸位置的速度进化,所以分子时钟可以在一个蛋白质中以不同的位置以不同的速度移动。蛋白质中的一些位点是保守的,这意味着这些序列位置上的氨基酸变化相对缓慢。进化的速度可能取决于物理性质,如残留物的位置(埋藏与暴露)或二级结构。例如,与埋藏或结构规则的区域(如螺旋和薄片)相比,暴露和无序的区域(如螺旋和线圈)经历了更快的替换。而具有许多构象灵活性的蛋白质区域往往进化得更快。这是因为这些区域可以在不破坏蛋白质功能的情况下容纳中性突变,而在埋藏或紧密排列的区域发生的突变则会破坏其结构的稳定,影响其功能。
定向进化是在实验室中改进蛋白质的一种方法
定向进化是在实验室中通过在试管中进行几轮突变和选择来修饰蛋白质的过程。假设你想修改一种蛋白质,使其更热稳定或提高催化活性。你从你感兴趣的天然蛋白质开始,然后你将突变随机地引入到该蛋白质的不同副本中。然后,你对感兴趣的特性进行分析或筛选,以发现哪些突变改善了蛋白质的特性。您选择这些改进的变体,放大它们,然后执行进一步的突变和选择图7.17)。这种方法的一个优点是,就像生物进化一样,它不需要基于知识的设计。
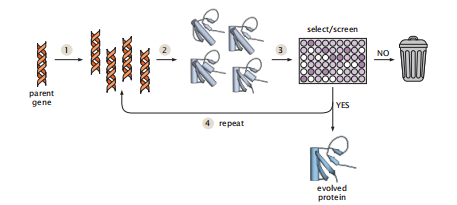
图7.17 定向进化导致了实验室中的新蛋白质。步骤1:随机突变亲本蛋白的基因,建立一个文库。步骤2:产生突变蛋白。步骤3:筛选蛋白质以选择所需的属性。拒绝那些没有显示出改善的突变体。步骤4:利用改良后的基因作为亲本,进行下一轮的突变和筛选。重复这个过程,直到进化出的蛋白质具有所需的特性。
定向进化在商业上用于生产具有优势工业特性的蛋白质,如增加表达水平、洗涤剂的酶活性、在不同温度或pH值下的稳定性、催化速率等。一些成功与定向进化的例子是增加1000倍的半衰期BPN没有Ca2+ ,增加1000倍的底物特异性β-半乳糖苷酶,105倍增加天冬氨酸转氨酶的活动β分支氨基酸和2-氧酸,和二聚氯酸突变酶转化为单体形式。
图7.18显示了一个细胞色素P450脂肪酸羟化酶通过9代突变和选择转化为丙烷单加氧酶的定向进化过程。进化出的酶在很大的温度范围内都具有良好的活性。这种进化导致了该酶的血红素结合袋附近的20个突变,重塑了底物进入途径。它说明了在非生物实验室的选择压力下,蛋白质如何被设计成具有期望的特性。定向进化也教会了我们关于自然进化的知识。以下是迄今为止所学到的两个关键经验教训。
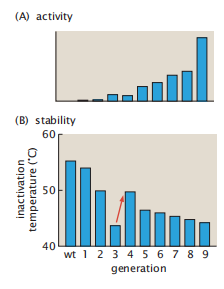
图7.18 定向进化系统地提供了更好的酶特性。(A)野生型细胞色素P450首先具有脂肪酸羟化酶活性。在9轮定向进化中,选择了丙烷单加氧酶(PMO)活性增加的突变体。(B)在前四轮试验中,蛋白质的稳定性下降。然后,选择蛋白质以获得更好的稳定性(红色箭头),然后活性提高。
首先,定向进化实验表明,随机突变和选择是穿越序列空间以达到蛋白质的期望特性的一个非常有效的组合。即使有益的变化相对较少,但几乎总是有良好的进化路线,包括中性或有益的突变,以实现定向进化的目标。这意味着蛋白质在变得更好的过程中很少需要产生适应性较差的状态。进化可以避免动力学陷阱,就像蛋白质折叠一样,因为被搜索的空间的维度是如此之大,所以总是有一些相对快速和直接的路线。
第二,定向进化实验表明,当蛋白质本身就是稳定的时候会提供丰富的进化路线。如果一种蛋白质是略微稳定的,那么许多可能的突变就会导致它的展开,而不是产生一个改进的蛋白质。这些进化路径都是死胡同。但如果一种蛋白质足够稳定,那么它就有更多的进化出口途径—更少的可能的突变会导致展开。请考虑下面的实验。从一个可以折叠成一个给定的天然结构的氨基酸序列开始。接下来,找到它的中性网络——所有与野生型序列不同的序列,至少有一个突变,仍然可以折叠成相同的结构。现在绕着这个逻辑走,找到具有最大突变稳定性的序列,也就是说,能够容忍最多的单个突变并仍然折叠到正确结构的序列。值得注意的是,在HP模型中发现,那些最高度连接的序列也往往具有最高的折叠稳定性。这意味着序列空间(如构象空间)具有漏斗状性质:蛋白质进化有利于具有最高中性空间连通性的蛋白质,而这些蛋白质也具有最大的构象稳定性。换句话说,不稳定蛋白质很少没有进化途径来获得更好的特性,因为大多数突变会展开不稳定蛋白质。相比之下,稳定蛋白有许多进化途径来获得更好的特性,因为稳定蛋白可以耐受一些不稳定的突变而不展开。
-总结
蛋白质序列通过突变和选择而进化。大多数单一突变不会改变蛋白质的结构或功能:天然结构对序列上的小变化是相当稳健的。但更广泛的突变可能导致蛋白质结构、动力学、聚集倾向、物理特性以及生物机制和功能的改变。进化的蛋白质会导致进化的生物体,有时会导致非常大的转化,比如新的生物物种。通常,如果两个蛋白质有相似的序列,它们也有相似的结构和功能。这种相关性是第8章中描述的方法的基础:通过检测氨基酸序列的相似性,你可以推断出蛋白质的结构和功能。
第七章完
-------------------------------------------
欢迎点赞收藏转发!
下次见!

















 792
792

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









