







表观遗传重编程是免疫细胞谱系指定的基础,但唯一定义免疫细胞类型的模式及其建立机制仍不清楚。在这里,我们从人类外周血中确定了六种免疫细胞类型的谱系特异性 DNA 甲基化特征,并确定了它们与其他表观遗传和转录组模式的关系。谱系特异性低甲基化的位点与每种细胞类型中不同的转录因子组合有关。相比之下,谱系特异性高甲基化的位点主要局限于适应性免疫细胞。PU.1 结合位点与不同细胞类型中的谱系特异性低甲基化和高甲基化有关,表明它以依赖于环境的方式调节 DNA 甲基化。这些观察结果表明,先天性和适应性免疫谱系由不同的表观遗传机制通过组合和依赖于环境的关键转录因子的使用来指定。所确定的细胞特异性表观基因组学和转录模式为未来研究疾病和衰老中的免疫失调奠定了基础。
简介
血液中的免疫细胞类型由骨髓中的造血干细胞 (HSC) 产生 (Dzierzak 和 Speck,2008;Mercier 等人,2011)。介导适应性免疫的 B 淋巴细胞和 T 淋巴细胞分别在骨髓和胸腺中发育。HSC 还产生先天免疫细胞,例如单核细胞、粒细胞和自然杀伤 (NK) 细胞。 免疫细胞的功能异质性是由通过表观遗传重编程选择性表达其相同遗传内容引起的。胞嘧啶残基处的 DNA 甲基化是哺乳动物中发现的最早的表观遗传修饰形式之一,受 DNA 甲基转移酶 (DNMT) 和十-十一-转位酶 (TET) 的作用调控 (Bestor 等人,2015 年;Ginno 等人,2020 年;Lio 和 Rao,2019 年)。作为表观遗传标记,甲基胞嘧啶修饰的化学稳定性及其双峰分布使其成为比较细胞状态(包括发育阶段或伴随疾病或衰老的变化)的有吸引力的候选对象。然而,唯一定义每种免疫细胞类型的甲基化特征及其建立的机制仍有待发现。
DNA 甲基化与造血细胞身份建立之间的关系在小鼠 B 淋巴细胞中得到了最全面的探索。小鼠的遗传研究表明,三种关键转录因子 E2A、Ebf1 和 Pax5 与此有关,在建立 B 细胞身份方面 (Hagman and Lukin, 2006; O’Riordan and Grosschedl, 1999)。在 E2A 的影响下,多潜能祖细胞进展到前原 B 细胞阶段时,DNA 甲基化变化明显 (Benner et al, 2015),并且随着 Ebf1 指导这些细胞启动抗原受体基因重排,进一步的变化随之产生 (Li et al, 2018)。在 B 细胞分化过程中,这些转录因子的结合基序在 Tet2 和 Tet3 蛋白诱导的差异甲基化区域 (DMR) 中富集 (Orlanski et al, 2016)。特定转录因子诱导甲基化变化的机制仍不清楚。

为了避免个体间差异并专注于生理相关的细胞类型特异性模式,这些模式与病理学的影响无关,我们绘制了 55 名健康个体的外周血中主要免疫细胞群的甲基化图谱,这些个体是根据严格的纳入标准招募的。我们确定并验证了六种细胞类型的谱系特异性甲基化特征,并将这些模式与其他表观遗传特征、基因表达和转录因子结合进行了比较。我们发现,细胞谱系特异性低甲基化位点和 DMR 与每种免疫细胞类型中转录因子结合位点的不同组合相关。相比之下,细胞特异性 DNA 高甲基化仅在适应性免疫细胞中突出。RNA 测序 (RNAseq) 进一步证实了适应性和先天性免疫细胞类型中 DNA 甲基化的差异使用。最后,PU.1 和 RUNX1 在不同细胞类型中以依赖于环境的方式与低甲基化和高甲基化相关。我们的研究揭示了在造血过程中建立的细胞特异性表观遗传和基因表达特征,并暗示一组有限的转录因子在定义人类免疫细胞身份方面发挥着作用。


结果
差异 DNA 甲基化可区分免疫细胞类型
为鉴定免疫细胞特异性 DNA 甲基化模式,我们使用磁珠和流式细胞术将人体血液分成 11 个亚组(图 1A 和 S1A;表 S1)。将每个样本的经亚硫酸氢钠处理的基因组 DNA 与 Illumina Infinium MethylationEPIC 阵列杂交,然后对每个探针进行甲基化处理和定量(b 范围从 0 到 1)(图 1B)。然后,我们确定了每种细胞类型中每个探针的平均 beta值。 得到的平均 beta值的归一化分布遵循特征性的双峰分布(图 1C)。第一个主成分 (PC1)(41%)将先天性(单核细胞、粒细胞和 NK 细胞)与适应性免疫亚组(B、CD4+ 和 CD8+ 淋巴细胞)区分开来。 PC2 (15%) 中骨髓来源的髓系细胞和 B 淋巴样细胞之间的额外分辨率明显 (图 1C)。幼稚 CD4+ 和 CD8+ T 细胞紧密聚集,在进一步的谱系特异性分析中,它们被视为一个区域。



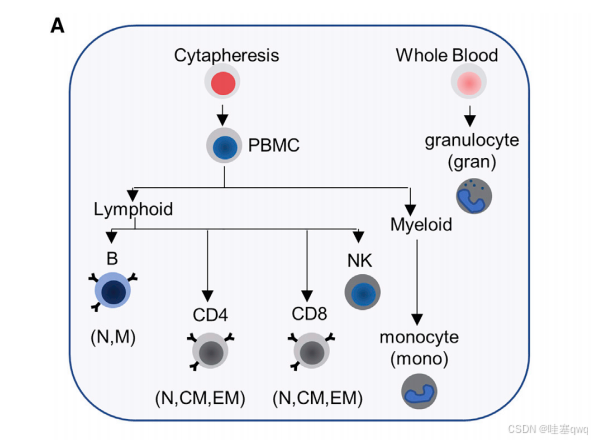
(A) 使用磁珠和流式细胞术从 55 名个体(年龄 22-83 岁)中纯化主要免疫细胞类型。亚硫酸氢钠处理的基因组 DNA 与 Illumina MethylationEPIC 阵列杂交
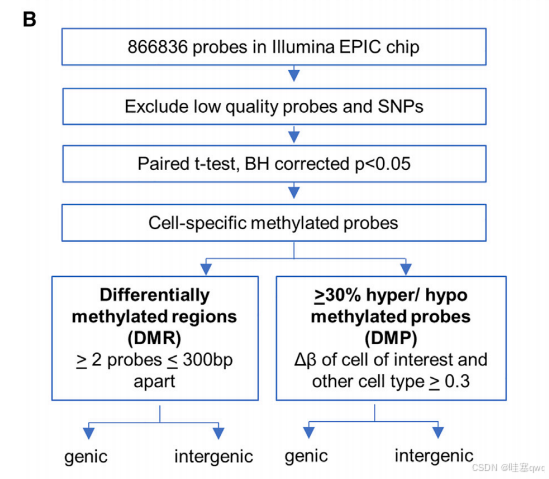
(B) 去除低质量探针后,对每种细胞类型进行配对 t 检验,比较所有参与者个体内 b 值与其他每种细胞类型的 b 值。在 BH 调整后的 p <0.05 下进行多重检验校正。细胞特异性低甲基化或高甲基化位点定义为某种细胞类型与所有其他细胞类型之间的 |Db| 为 R0.3 的位点。所有进一步分析均使用个体间共享的细胞特异性差异甲基化探针。根据 300 bp 内两个或更多差异甲基化探针的存在来确定差异甲基化区域(表 S3)。

 最低0.47元/天 解锁文章
最低0.47元/天 解锁文章


















 1243
1243

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









