




nee2024 经历高速范围变化的海洋鱼类可能不是气候变化的赢家 Marine fishes experiencing high-velocity range shifts may not be climate change winners 论文图像内容与结论总结(学习记录)
一、图像内容及支撑结论
1. 图1:“迁移情景”下种群丰度趋势与范围迁移速率的理论关联图
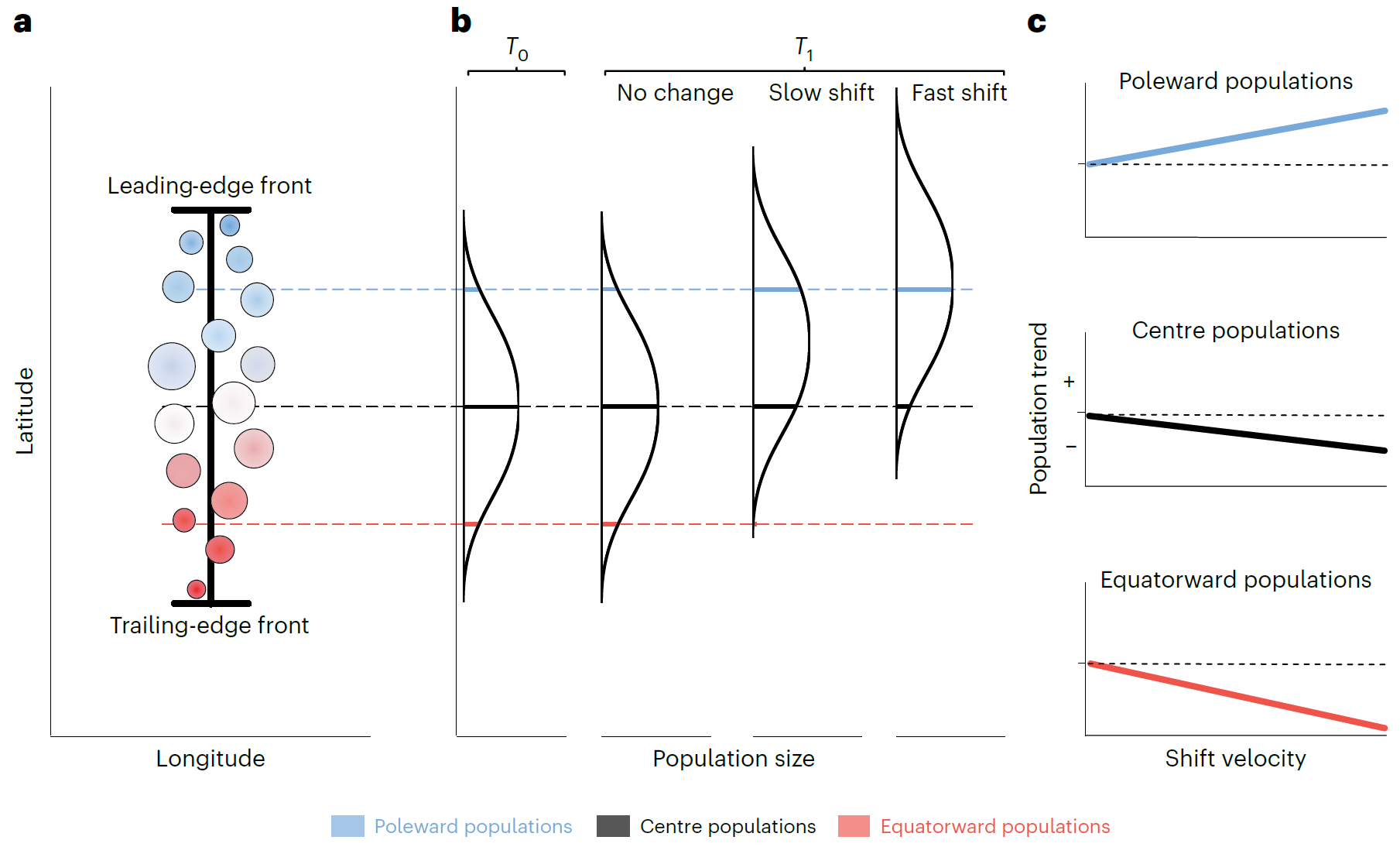
- 图像内容:包含3个子图。a图为物种纬度范围内种群分布理论图,圆点代表不同种群,点大小与种群规模正相关,颜色区分种群到极向(蓝色)和赤道向(红色)边缘的距离,黑色线及误差线表示纬度范围,虚线标注极向、中心、赤道向种群位置;b图为基线条件(左,T₀)与三种迁移速率情景(右,T₁)下种群规模与空间位置的关系,单峰曲线代表全范围丰度分布,彩色虚线标注三类种群,粗线突出不同迁移速率下种群规模相对于T₀的变化;c图为b图情景下种群丰度趋势与迁移速率的预期关系,显示“迁移情景”下极向种群随迁移速率升高丰度增加,赤道向种群减少。
- 支撑结论:提出“迁移情景”的理论假设——若物种总丰度稳定,极向迁移物种应呈现“极向种群增加、赤道向种群减少”的对称变化;为后续实证分析提供理论对照,凸显实际观测与理论预期的差异。
2. 图2:范围迁移速率对种群趋势的影响图
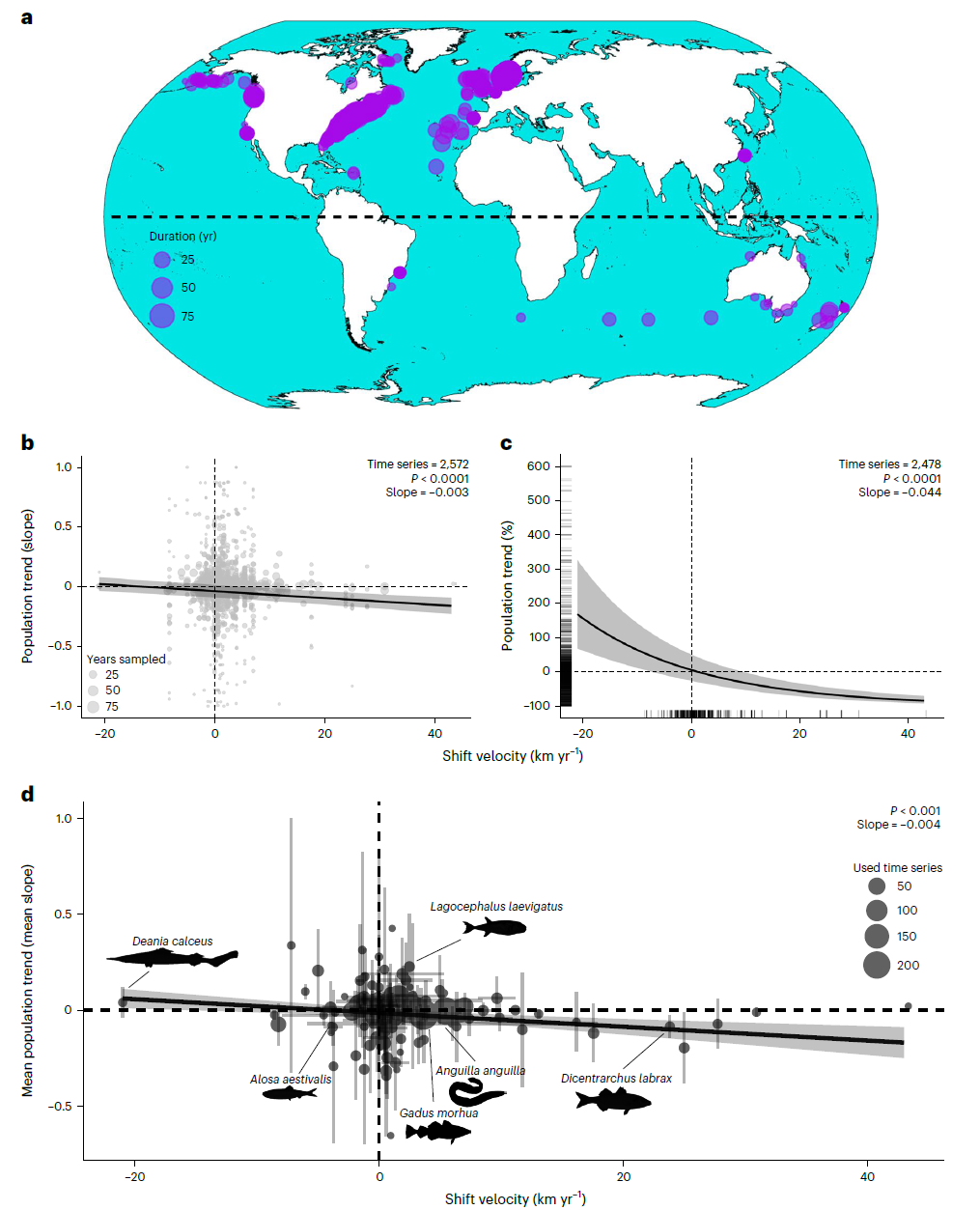
- 图像内容:包含4个子图。a图为采样种群的空间分布图(罗宾逊投影),圆点代表种群时间序列,点大小与时间序列持续时间(年)正相关,覆盖大西洋、太平洋、印度洋等全球海域;b图为种群趋势(标准化丰度对采样年份的回归斜率)与质心迁移速率的关系,圆点为单个种群(点大小代表采样年数),黑色线为GLMM模型斜率(-0.003,P<0.0001),灰色区域为95%置信区间,显示迁移速率越高,种群趋势越负;c图为种群趋势百分比变化(非标准化,泊松回归估算)与质心迁移速率的关系,y轴为T₁₀与T₁的丰度比, rug线标注数据点,移除>700%的极端值后,17km/年(99百分位)迁移速率对应10年丰度下降50%,非迁移物种(25百分位)丰度增加5.9%;d图为物种水平平均种群趋势与平均质心迁移速率的关系(n=146种),x轴正值为极向迁移,虚线为零变化线,误差线为95%置信区间,显示高速率极向迁移物种的平均种群趋势为负。
- 支撑结论:实证证实极向迁移速率与种群趋势负相关,高速率迁移(17km/年)导致种群显著衰退,且该关联在物种水平依然成立;颠覆“快速迁移物种是气候适应赢家”的假设,为“迁移与崩溃”情景提供核心数据支撑。
3. 图3:种群空间位置与热位置对迁移-种群趋势关联的调节图
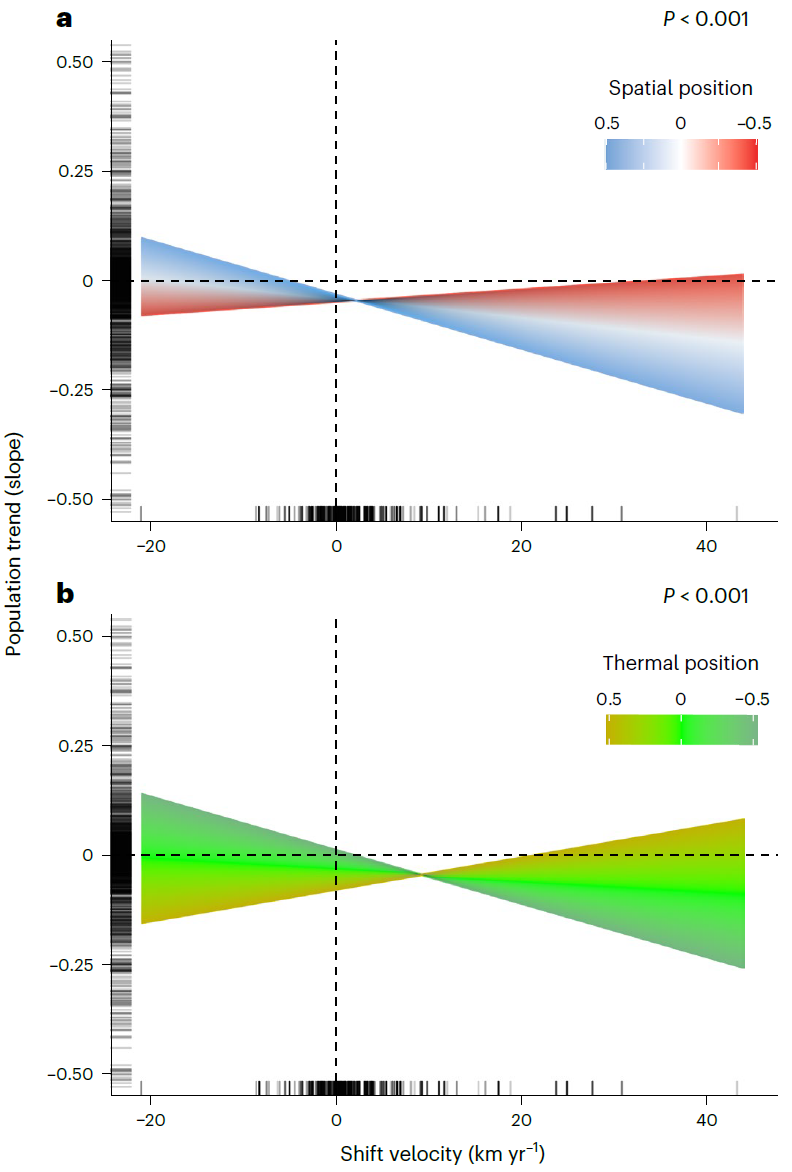
- 图像内容:包含2个子图,均基于GLMM模型,y轴为种群趋势(斜率),x轴为迁移速率(极向为正),rug线标注数据点。a图为空间位置(种群相对于物种范围的位置)与迁移速率的交互效应,蓝色正值代表极向种群,红色负值代表赤道向种群,显示极向种群随迁移速率升高趋势更负(P<0.001),赤道向种群负趋势更弱;b图为热位置(种群相对于物种热偏好的位置)与迁移速率的交互效应,橙色正值代表种群所处环境高于物种热偏好(暖边缘),浅绿色负值代表低于热偏好(冷边缘),显示冷边缘种群随迁移速率升高趋势更负(P<0.001),暖边缘种群趋势更平缓。
- 支撑结论:明确种群在物种范围和热偏好中的位置调节迁移速率的效应——极向、冷边缘种群对迁移速率更敏感,衰退更显著;赤道向、暖边缘种群受影响较小,进一步细化“迁移与崩溃”情景的种群差异。
4. 图4:与实证结果一致的极向偏斜丰度分布示意图
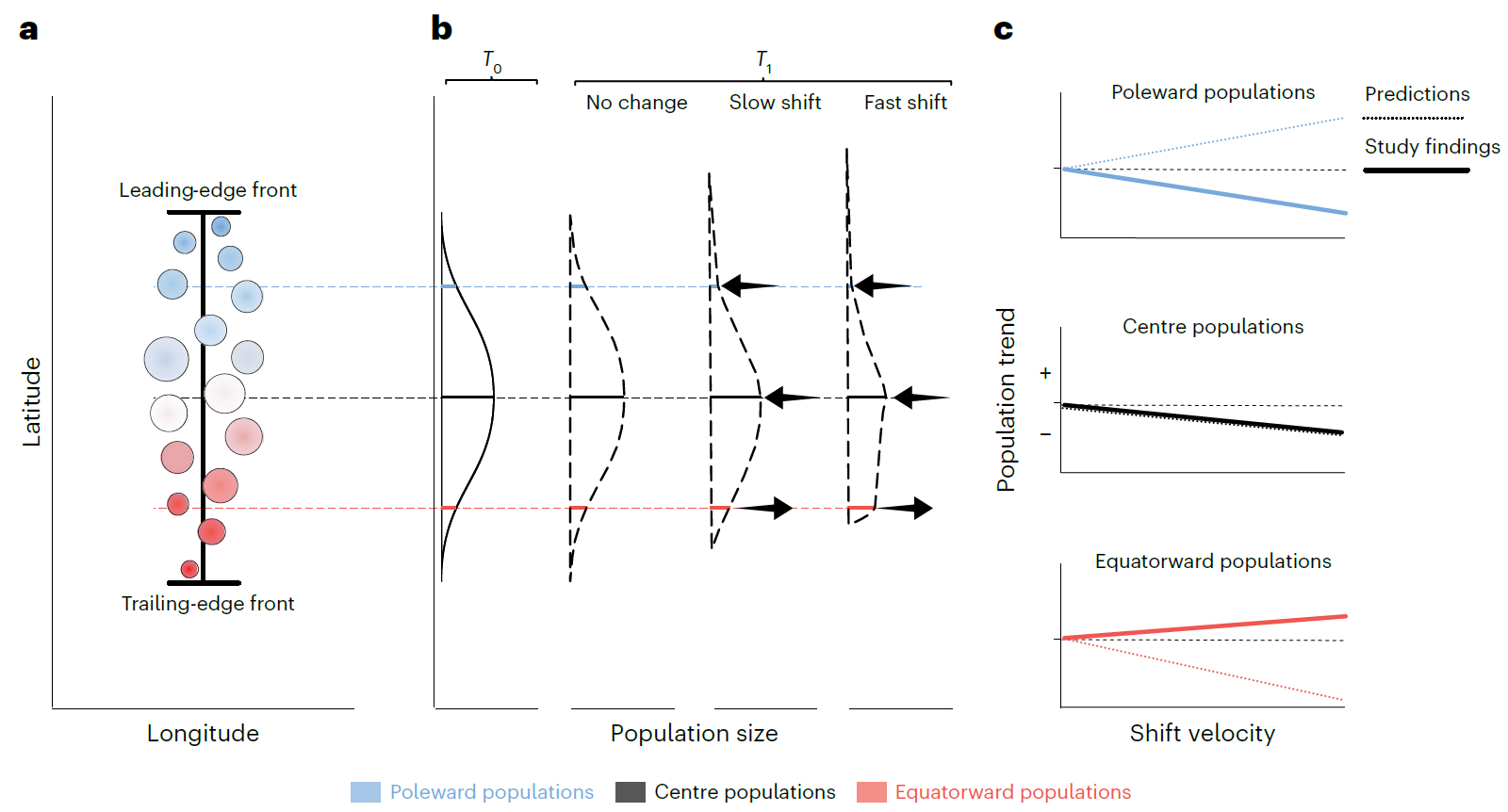
- 图像内容:结构与图1对应,共3个子图。a图同图1a,为物种纬度范围内种群分布理论图;b图为基线条件(左,T₀)与三种迁移速率建议(右,T₁)下种群规模与空间位置的关系,曲线代表全范围丰度分布,黑色箭头标注各类种群丰度变化方向(左为减少,右为增加),显示极向种群随迁移速率升高丰度减少,赤道向种群稳定或略增;c图为b图情景下种群趋势与迁移速率的关系(实线),与图1c的理论预期(虚线)对比,凸显实际观测中极向种群随迁移速率升高趋势下降的特征。
- 支撑结论:提出“极向偏斜丰度分布”的修正模型——快速迁移物种的丰度分布呈非对称变化,极向种群衰退、赤道向种群稳定,解释实证观测与“迁移情景”理论的差异,为“迁移与崩溃”情景提供理论支撑。
二、论文核心结论
1. 极向范围迁移与种群趋势负相关,颠覆“气候适应赢家”假设
- 迁移速率的负面影响:全球2572个海洋鱼类种群(146种)的实证数据显示,极向迁移速率与种群趋势显著负相关(GLMM:slope=-0.003,P<0.0001)。高速率迁移(17km/年,99百分位)的种群,10年内丰度平均下降50%(95%CI:25.6%66.6%),而非迁移物种(25百分位)丰度仅增加5.9%(95%CI:-25.8%51.07%),如Trisopterus luscus(17.6km/年迁移)丰度下降92.7%,Syngnathus acus(25km/年迁移)下降75.5%🔶11-55、11-73。
- 物种水平一致性:物种水平平均种群趋势与平均迁移速率同样呈负相关(GLMM:slope=-0.004,P<0.001),排除“少数物种驱动结果”的干扰,证实该关联具有跨物种普遍性🔶11-74。
2. 极向、冷边缘种群对迁移速率更敏感,衰退更显著
- 空间位置调节效应:种群在物种范围内的位置显著调节迁移速率的影响(GLMM:交互效应z=-3.485,P<0.001)。极向种群随迁移速率升高,负趋势更显著;赤道向种群负趋势较弱;中心种群呈轻微负趋势,与“迁移情景”中“极向种群增加”的理论预期完全相反🔶11-82。
- 热位置调节效应:种群相对于物种热偏好的位置同样产生调节(GLMM:交互效应z=3.412,P<0.001)。冷边缘种群(环境温度低于物种热中点)随迁移速率升高衰退更显著;暖边缘种群(环境温度高于热中点)趋势更平缓,说明热适应性差异加剧种群对迁移的响应分化🔶11-83。
3. 捕捞压力、纬度等因素不主导迁移-种群趋势关联
- 捕捞压力的有限影响:尽管高商业价值物种整体衰退更显著(GLMM:效应量=0.029,P<0.001),但捕捞脆弱性与迁移速率无显著交互作用(GLMM:z=-0.814,P=0.415),且低捕捞脆弱性物种仍呈现“极向种群随迁移速率衰退”的核心模式,说明气候驱动的迁移是种群衰退的重要因素,而非捕捞主导🔶11-90、11-91。
- 纬度无直接影响:种群趋势与绝对纬度无显著关联(补充图7a),且包含迁移速率的模型AIC显著低于包含纬度的模型(ΔAIC>24),证实迁移速率对丰度变化的影响强于纬度,排除“高纬度升温快导致种群衰退”的简单解释🔶11-88。
4. 快速迁移物种呈现“极向偏斜丰度分布”,面临气候脆弱性
- 非对称丰度变化:快速迁移物种的丰度分布呈“极向种群衰退、赤道向种群稳定”的非对称变化(图4),与“迁移情景”的对称预期不符,可能源于极向种群面临更高的气候变异(如极端温度事件)和突变负荷,导致局部丰度下降🔶11-101、11-102。
- 保护启示:快速极向迁移的海洋鱼类(尤其是极向、冷边缘种群)并非“气候赢家”,反而属于“气候输家”,需纳入高保护优先级;现有保护应超越“仅关注范围边缘”,覆盖全范围种群变化,避免低估气候对物种存续的威胁🔶11-105。
三、论文研究方法总结(基于s41559-024-02350-7.pdf)
3.1 核心数据来源
本研究通过整合两大全球数据库(BioTIME、BioShifts)及多源辅助数据,构建“种群趋势-范围迁移速率”关联分析的数据集,具体来源与用途如下:
3.1.1 种群趋势数据:BioTIME数据库
- 数据库定位:全球最大的生物多样性时间序列数据库,涵盖319项独立研究的物种丰度或出现数据,可量化不同时空尺度的种群动态变化🔶12-101、12-177。
- 数据筛选与标准化:
- 种群定义:将“同一研究+同一BioTIME‘plot’(重复采样位点)+同一物种”的丰度记录定义为1个种群,若种群内年度采样位点纬度差异>1°,则通过坐标平均剔除异常位点,确保单一种群的空间一致性(最终种群纬度差异均值±标准差为0.005°±0.05°)🔶12-105。
- 质量控制:
- 剔除“采样次数<3次”或“全时间序列丰度为0”的种群,避免抽样偏差;
- 对1年内多次采样的种群(n=793个),计算年均丰度以统一时间分辨率;
- 排除“物种数<5或个体数<10”的时间序列,确保群落数据的代表性🔶12-105。
- 最终数据集:提取2572个地理编码的种群级丰度变化数据,覆盖146种海洋鱼类(68科),时间序列持续时间均值±标准差为34±26.9年(中位数26年),采样年数均值±标准差为17.05±18.2年(中位数9年),涵盖温带、亚极区物种及少量热带物种,分布于大西洋、太平洋、印度洋等全球海域🔶12-105、12-106。
3.1.2 范围迁移速率数据:BioShifts数据库
- 数据库定位:全球气候驱动物种再分布地理数据库,整合文献中报道的范围迁移速率估算结果,涵盖陆地与海洋生态系统🔶12-109、12-171。
- 数据筛选与关键参数:
- 迁移速率定义:以“纬度范围迁移速率(km/年)”为核心指标,正值代表极向迁移,负值代表赤道向迁移;每个估算值均基于至少2次历史调查(间隔≥10年),并标注迁移发生的范围位置(质心、极向边缘、赤道向边缘)🔶12-109。
- 数据选择:
- 优先选择“范围质心迁移速率”数据(占物种总数的81%),因质心可反映物种整体分布的移动趋势,避免边缘迁移的局部偏差;
- 最终匹配146种鱼类的迁移速率数据,速率范围为-21~+43.4km/年,均值±标准误为1.96±0.08km/年(整体呈极向迁移趋势)🔶12-109、12-42。
3.1.3 辅助数据:空间位置、热位置与捕捞压力
| 数据类型 | 来源与计算方法 | 核心用途 |
|---|---|---|
| 物种分布范围与空间位置 | 物种分布边界从GBIF数据库提取,经CoordinateCleaner包过滤(剔除重复记录、陆地记录、机构标本记录等);通过0.1/0.9分位数估算纬度范围,计算“种群-范围中心”的纬度距离占比,得到标准化空间位置指数(正值=极向侧,负值=赤道向侧)🔶12-115、12-116 | 量化种群在物种分布范围内的相对位置,分析空间位置对迁移-种群趋势关联的调节作用 |
| 热位置与海表温度 | 物种热偏好数据来自Bio-ORACLE数据库(2000-2014年月均海表温度,分辨率0.083°),通过空间 thinning(0.083°分辨率)平衡采样密度;计算“种群温度-物种热中点”的差值占比,得到标准化热位置指数(正值=暖边缘,负值=冷边缘)🔶12-122、12-123 | 量化种群相对于物种热偏好的位置,分析热适应性对迁移响应的影响 |
| 捕捞压力指数 | 捕捞脆弱性指数来自Cheung等(2005)的研究,基于繁殖力、寿命、性成熟年龄等生活史特征计算(1~100分,分值越高越脆弱);商业状态基于FishBase分类,合并为“商业种”(高/中商业价值)与“非商业种”(低/无商业价值)两类🔶12-128、12-129 | 控制捕捞压力对种群趋势的干扰,验证气候迁移的独立影响 |
3.2 数据处理关键流程
3.2.1 分类学匹配与时空匹配
- 分类学统一:通过GBIF Backbone Taxonomy工具(rgbif R包)匹配BioTIME与BioShifts的物种学名,整合World Register of Marine Species等权威分类数据源,解决异名问题,确保146种鱼类在两大数据库中的分类一致性🔶12-110、12-209。
- 时空不匹配校正:
- 空间不匹配:计算BioTIME种群位点与BioShifts迁移估算位点的大圆距离(空间不匹配度),后续通过统计模型纳入该变量,检验“空间距离越近,迁移-种群趋势关联越强”的假设🔶12-113。
- 时间不匹配:计算BioTIME种群首次采样年份与BioShifts迁移估算首次年份的差值(时间不匹配度),验证“迁移估算与种群采样时间重叠时,关联更显著”的预期🔶12-113、12-61。
3.2.2 种群趋势量化
采用两种丰度指标计算种群趋势,兼顾数据标准化与生物学解释性:
- 标准化丰度(用于模型拟合):对每个种群的丰度数据进行“均值减-标准差除”标准化,消除不同种群丰度量级差异;通过线性模型以“标准化丰度”为响应变量、“采样年份”为预测变量,将回归斜率定义为“种群趋势”(正值=丰度增加,负值=丰度减少)🔶12-107。
- 非标准化丰度(用于结果解释):对原始丰度数据(排除出现/密度型时间序列,n=94个)构建泊松回归模型,估算“时间序列第10年(T₁₀)与第1年(T₁)的丰度比”,转化为百分比变化(如50%下降表示T₁₀丰度为T₁的50%),增强结果的生物学意义🔶12-107。
3.2.3 异常值与敏感性处理
- 极端值控制:
- 迁移速率:通过“1.5倍四分位距”筛选,排除极端迁移速率值(>1.5×IQR),验证核心结论是否受极端值驱动;
- 种群趋势:在百分比变化分析中,移除>700%的极端值(n=122个),避免少数异常种群干扰整体趋势🔶12-35、12-48。
- 多情景验证:
- 迁移速率类型:分别采用“最近质心迁移速率”与“物种平均迁移速率”,结果一致;
- 数据质量:区分BioShifts中“实际数据”(86.4%)与“重测数据”(13.6%),验证不同数据质量对结果的影响;
- 种群趋势可靠性:仅保留“线性模型R²≥0.5”的种群(n=328个),核心负相关关联仍显著(GLMM:slope=-0.017,P<0.001)🔶12-48、12-112、12-136。
3.3 统计分析方法
3.3.1 核心模型:广义线性混合效应模型(GLMM)
- 模型框架:采用glmmTMB R包构建GLMM,以“种群趋势”为响应变量,“范围迁移速率”为核心预测变量,控制以下随机效应与权重:
- 随机效应:纳入“分类学科”与“BioTIME研究ID”作为随机截距,解释科间差异与研究方法的系统误差(未纳入物种随机效应,因多数物种的种群样本量少)🔶12-134、12-135;
- 权重:以“种群采样次数的平方根”为权重,降低低可靠性(采样次数少)种群的影响;敏感性分析显示,采用“1/种群斜率标准误”作为权重,结果无定性差异🔶12-135。
- 关键交互模型:
- 空间位置调节模型:纳入“迁移速率×空间位置”交互项,检验种群在物种范围内的位置对关联的调节;
- 热位置调节模型:纳入“迁移速率×热位置”交互项,区分暖边缘与冷边缘种群的响应差异;
- 捕捞压力交互模型:纳入“迁移速率×捕捞脆弱性”“迁移速率×空间位置×捕捞脆弱性”交互项,控制捕捞的潜在干扰🔶12-139、12-147。
3.3.2 模型验证与诊断
- 假设检验:通过残差正态性检验、方差膨胀因子(VIF≤3)检验、同方差性检验,确保模型满足GLMM假设;
- 边际R²计算:采用MuMIn R包的r.squaredGLMM函数,区分“固定效应解释的方差(边际R²)”与“固定+随机效应解释的方差(条件R²)”,明确迁移速率等固定效应的实际解释力(如核心模型边际R²=0.003,条件R²=0.62,反映种群趋势的异质性高)🔶12-138、12-224;
- 区域与栖息地子集分析:将全球海域分为东北大西洋、西北大西洋、东北太平洋等5个区域,将鱼类分为底栖(85种,1464个种群)、中上层(27种,324个种群)等类群,验证核心关联的区域与栖息地一致性🔶12-146、12-127。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









