



本文内容速览:
在之前的文章“植物微生物组与植物抗病”中,小远给大家介绍了植物生长环境中各种微生物对植物产生的危害,其中细菌是导致农业作物病害的一大真凶。你家阳台种的番茄是不是突然蔫了?叶片发黄、茎秆一捏就软,拔起来看根部发黑发黏 —— 这大概率是细菌性青枯病在作祟。这种由青枯雷尔氏菌(Ralstonia solanacearum)引起的病害,能让番茄、烟草、辣椒等200多种植物减产30%~100%。更严重的是,传统铜制剂、抗生素农药的过度使用,不仅会使土壤的重金属含量超标,还会导致细菌进化出各种抗性,变成超级细菌。在最近的研究中,科学家们发现出了一个“天然解决方案”——噬菌体(细菌病毒)。这种比细菌小100倍的“微型杀手”,特异性针对有害细菌,对植物、蜜蜂、人类等都无害,甚至能和农药协作提升效果。今天就以青枯雷尔氏菌为例,跟小远一起看看噬菌体是怎么成为植物的天然卫士,以及它从实验室走向田间,还克服了哪些难题。
一、关于噬菌体
提到“病毒”,大家可能会想到流感、新冠,但噬菌体是只侵染细菌的特殊病毒。它们是地球上数量最多的生物 —— 每克土壤里就有10亿个噬菌体,比全地球人口还多。
1. 噬菌体的两种生命周期
噬菌体的一生只做一件事:找到并杀死特定细菌。根据噬菌体的生命活动周期,大致分为两种类型(Halawa, 2023):
· 溶菌性噬菌体(烈性噬菌体):“快准狠”型杀手。先通过尾部纤维识别与吸附细菌表面的脂多糖、鞭毛等结构蛋白,然后把自己的DNA“注射”进细菌体内,利用细菌的酶、原料和核糖体,复制自身遗传物质并合成外壳蛋白,批量制造新噬菌体,最后释放溶菌酶杀死细菌,并释放出几百个后代继续“追杀”其他细菌。现在用来防治植物细菌病害的,基本上全是这种只杀死细菌并且无法整合到细菌基因组上的噬菌体。
· 溶原性噬菌体(温和噬菌体):“潜伏者”型杀手。将DNA插入细菌基因组,跟着细菌分裂一起复制,暂时不杀死宿主细菌;但当遇到紫外线、农药等刺激,就会从细菌基因组上切除,变成溶菌性噬菌体,杀死细菌。这种噬菌体在农业防治上有一定风险——比如有的会产生一些毒力因子,让致病菌毒力更强,所以在农业上基本上不会使用这类噬菌体。
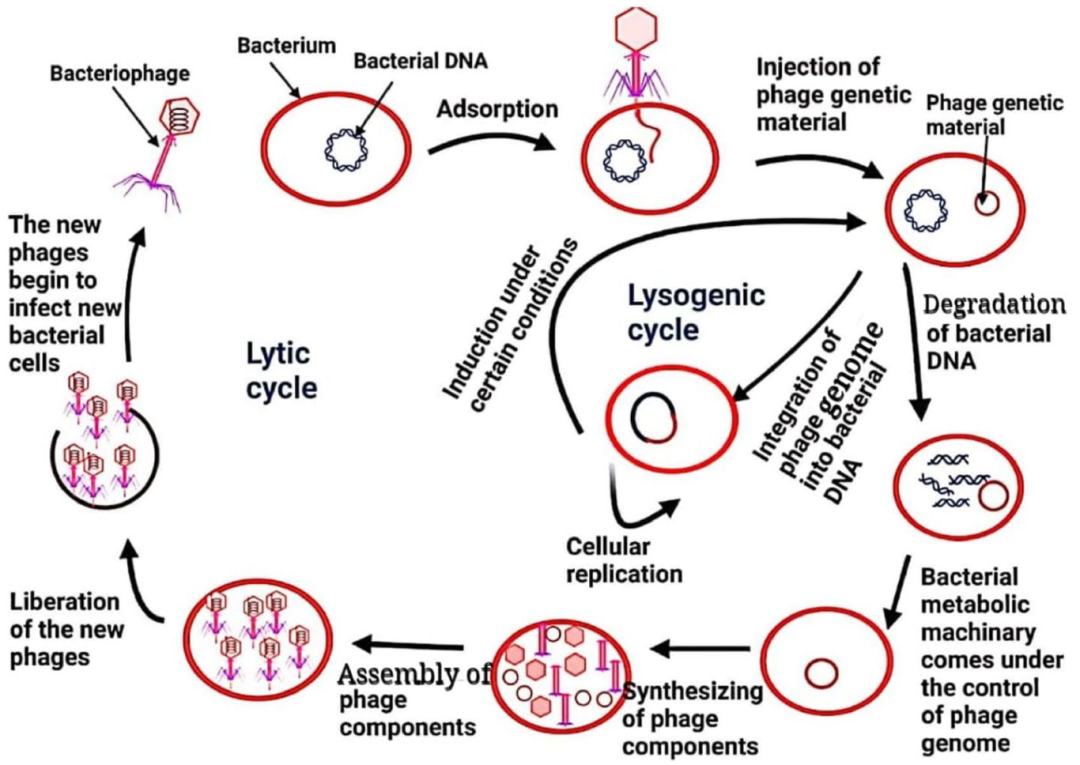
图1 噬菌体感染细菌的两种路径(Halawa, 2023)。溶菌性路径(右)直接裂解细菌释放子代;溶原性路径(左)则将 DNA 整合进细菌基因组,等待激活信号。
2. 噬菌体适用于农业的特征
· 绝对专一:只杀目标细菌,比如青枯雷尔氏菌噬菌体不会侵染根瘤菌(帮助豆科植物固氮的有益细菌),也不会伤害蜜蜂、蚯蚓等昆虫(Halawa, 2023);
· 自我增殖:只要有宿主细菌,噬菌体就可以不断繁殖,不用反复喷施;
· 无残留:噬菌体由蛋白质和DNA组成,会被土壤微生物分解,不会像铜制剂等化学药品一样在土壤里积累;
· 不怕抗性:随着细菌进化出抗性,噬菌体也可能会共同进化,甚至能让抗性细菌毒性变弱——比如青枯雷尔氏菌为了抵抗噬菌体,会减少用来入侵植物的鞭毛、胞外多糖等物质的合成,从而导致毒性下降。
二、噬菌体应用的真实案例
近年来,科学家已经分离出上百种针对植物病原菌的噬菌体,其中针对青枯雷尔氏菌、番茄斑点病菌的噬菌体,已经在田间试验中实现80%的病害减少。在此,小远以两个代表性的噬菌体为例,看看它们是怎么抵抗病害的。
案例 1:LPRS20—— 青枯病的 “全能杀手”
2024年10月,华中农业大学李锦铨教授课题组和广东省农科院农业农村部南方植物营养与肥料重点实验室在Journal of Agricultural and Food Chemistry期刊上发表了一篇题为“Exploration of Phage−Agrochemical Interaction Based on a Novel Potent Phage LPRS20-Targeting Ralstonia solanacearum”的研究论文,作者从广东烟草田间土壤里,分离出一种新型噬菌体LPRS20。它不仅是首次被发现的Ampunavirus 属新物种,还是个 “全能选手”:
1. 清除细菌:广谱且高效
青枯雷尔氏菌具有多样性,不同作物上的菌株差异很大,有的噬菌体只能靶向番茄上的菌株,对烟草上的菌株却没用。但LPRS20不一样——它能裂解7种青枯雷尔氏菌,包括感染烟草的LRS3、番茄的LRS2、辣椒的LRS1。
在生长曲线实验中,LPRS20表现出的杀菌效率更是惊人:当噬菌体和青枯雷尔氏菌的比例(MOI)达到0.1时,24小时就能让细菌数量减少2个数量级;如果MOI提升到100,几乎能让细菌无法生长。对比传统农药小檗碱(Berberine),LPRS20的抑菌效果更持久,48小时后细菌仍没恢复生长。
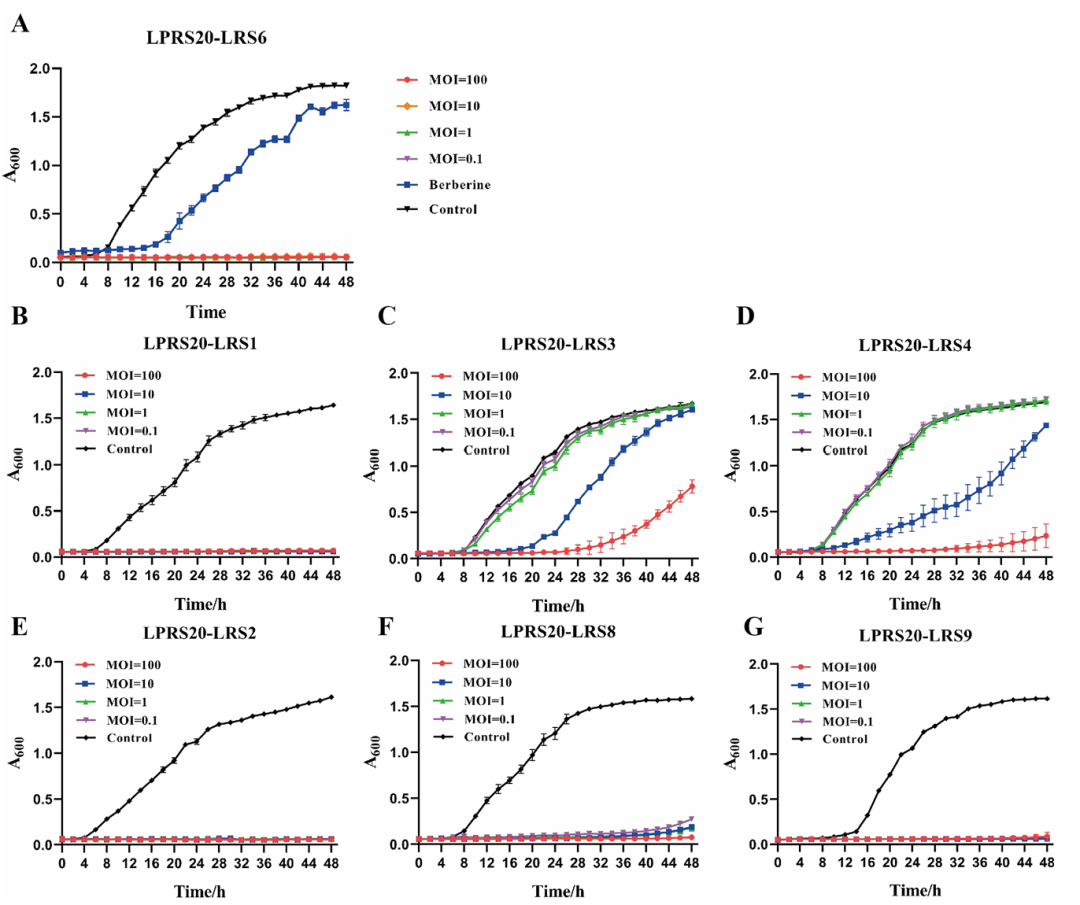
图2 LPRS20对7种青枯雷尔氏菌的生长抑制曲线(Liu et al., 2024)。(A)使用烟草上的菌株LRS6,MOI=100时,48小时内OD600(细菌密度指标)接近 0,远低于对照组和小檗碱组;(B-G)使用辣椒、番茄上的菌株时,均表现出显著抑制。
2. 面对植物:绝对安全
很多农药的使用会让植物产生“药害”——比如高浓度铜制剂会让烟草叶片出现斑点、发黄的症状。但LPRS20对植物几乎没有伤害:作者用1010 PFU/mL(超高浓度)的LPRS20喷洒烟草幼苗,72小时后观察叶片,既没有斑点、卷曲,也没有枯萎。通过感官评估,可以看出噬菌体处理组的叶片健康度和喷PBS缓冲液的对照组几乎没差别,而噻唑锌铜(Thiodiazole-copper)处理组的叶片已经出现明显斑点(图3)。

图3 72小时后不同处理的烟草叶片(Liu et al., 2024)。第一行为LPRS20(5~10 log₁₀ PFU/mL),叶片无任何药害;中间一行为噻唑锌铜(16~128 μg/mL),高浓度组出现斑点和卷曲的现象;第三行为小檗碱(8~128 μg/mL),高浓度组出现轻微卷曲的现象。
3. 土壤环境:保持活性
植物病害的病原菌有时会藏在土壤里,而一些农药有时不仅难以渗入土壤甚至还会降低土壤质量,但噬菌体可以解决这个问题。在灭菌土壤中加入青枯雷尔氏菌后,加入LPRS20,24小时后细菌数量显著减少,比小檗碱的效果强1倍(图4A)。
更关键的是,在田间试验中,烟草感染青枯雷尔氏菌1天后喷洒LPRS20,22天后烟草的病害指数从95.06(对照组)降到51.85,防治效率达到45.46%(图4B、C)。
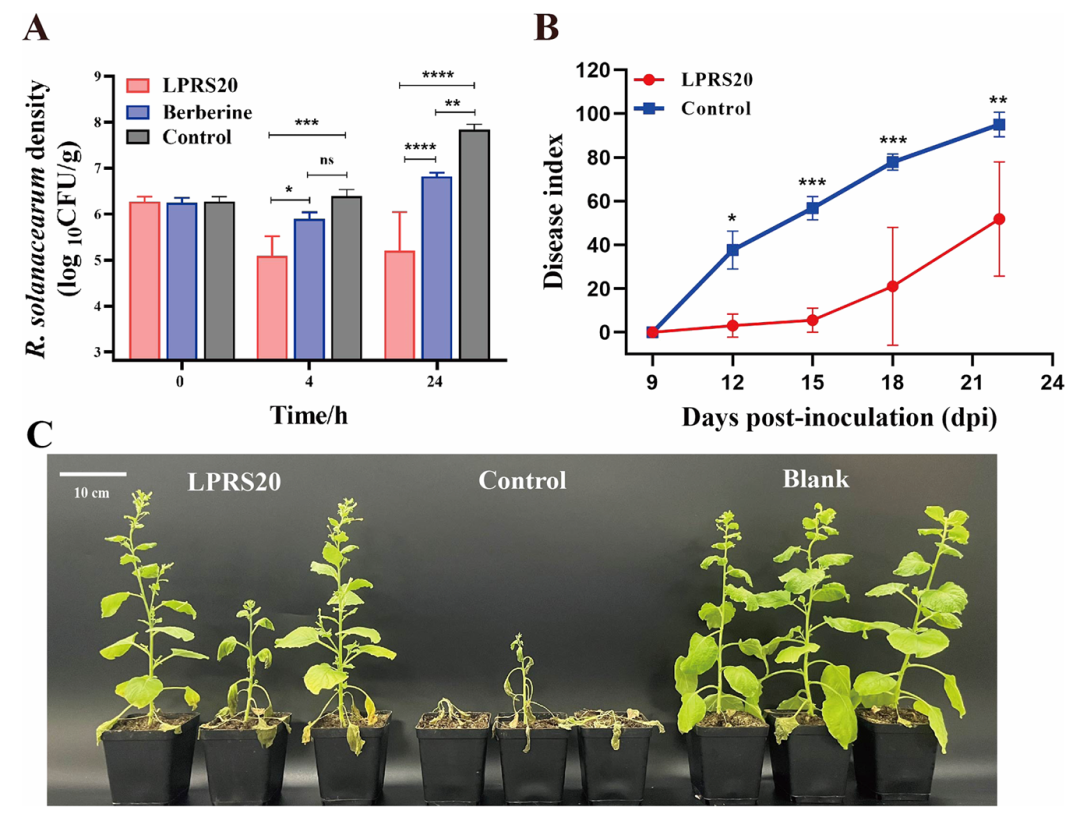
图4 土壤和烟草中噬菌体LPRS20的生物防治试验(Liu et al., 2024)。(A)土壤中 LPRS20对青枯雷尔氏菌的抑制情况(24小时减少2.64 log₁₀ CFU/g);(B)烟草病害指数变化情况(22天防治效率45.46%);(C)不同处理下,22天后的烟草植株,对照组(中间)几乎枯萎,LPRS20处理组(右)长势接近空白组(左)。
案例 2:噬菌体组合 ——1+1>2的超强防治能力
如果在治理病害的过程中只使用单一噬菌体,细菌可能会进化出抗性,但研究人员发现了应对办法——噬菌体组合疗法。
1. 多种噬菌体协同使用
2019年12月,南京农业大学资源与环境科学学院沈其荣课题组在Nature Biotechnology杂志上发表了一篇题为“Phage combination therapies for bacterial wilt disease in tomato”的研究论文,他们从番茄田分离出了4种青枯雷尔氏菌噬菌体(NJ-P3、NB-P21、NC-P34和NN-P42)。在单独使用其中一种噬菌体时,番茄青枯病发病率能降低到40%;但把4种噬菌体混合使用,发病率直接降到20%以下,田间病害指数减少80%(图5)。
该治疗方法效率更高有两个关键原因:
· 减少抗性:细菌要同时抵抗4种噬菌体,需要突变4个不同的基因,难度比抵抗1种高100倍;
· “弱化”细菌:就算有少数细菌进化出抗性,也会付出代价——比如为了不被噬菌体识别,它们会减少鞭毛或胞外多糖的合成,而这些正是用来入侵植物的关键因子,结果导致生长速度变慢,毒力下降(图6)。实验发现,抗性菌株的生长能力(OD600)比普通菌株低30%,在土壤里难以跟其它微生物竞争。
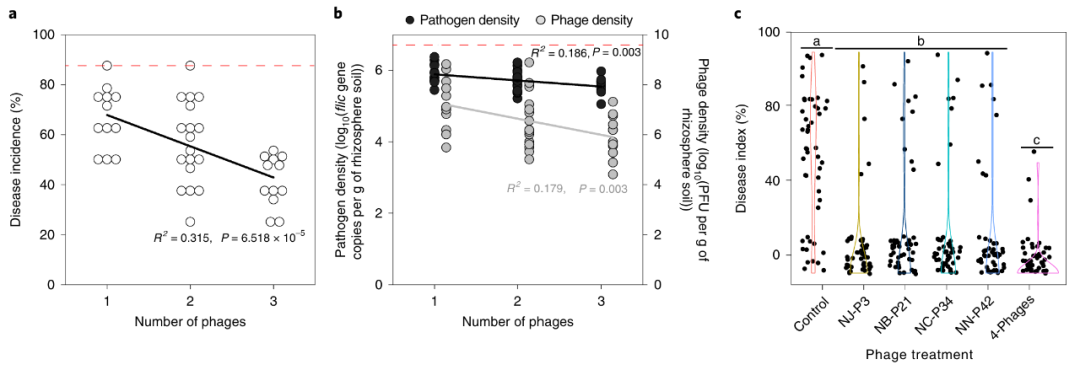
图5 噬菌体组合以及病害发生率(Wang et al., 2019)。(A)温室实验中,噬菌体种类越多,番茄青枯病发病率越低(R²=0.315,P<0.001);(B)病原菌密度随噬菌体数量增加而减少;(C)田间试验,4种噬菌体组合的病害指数最低。
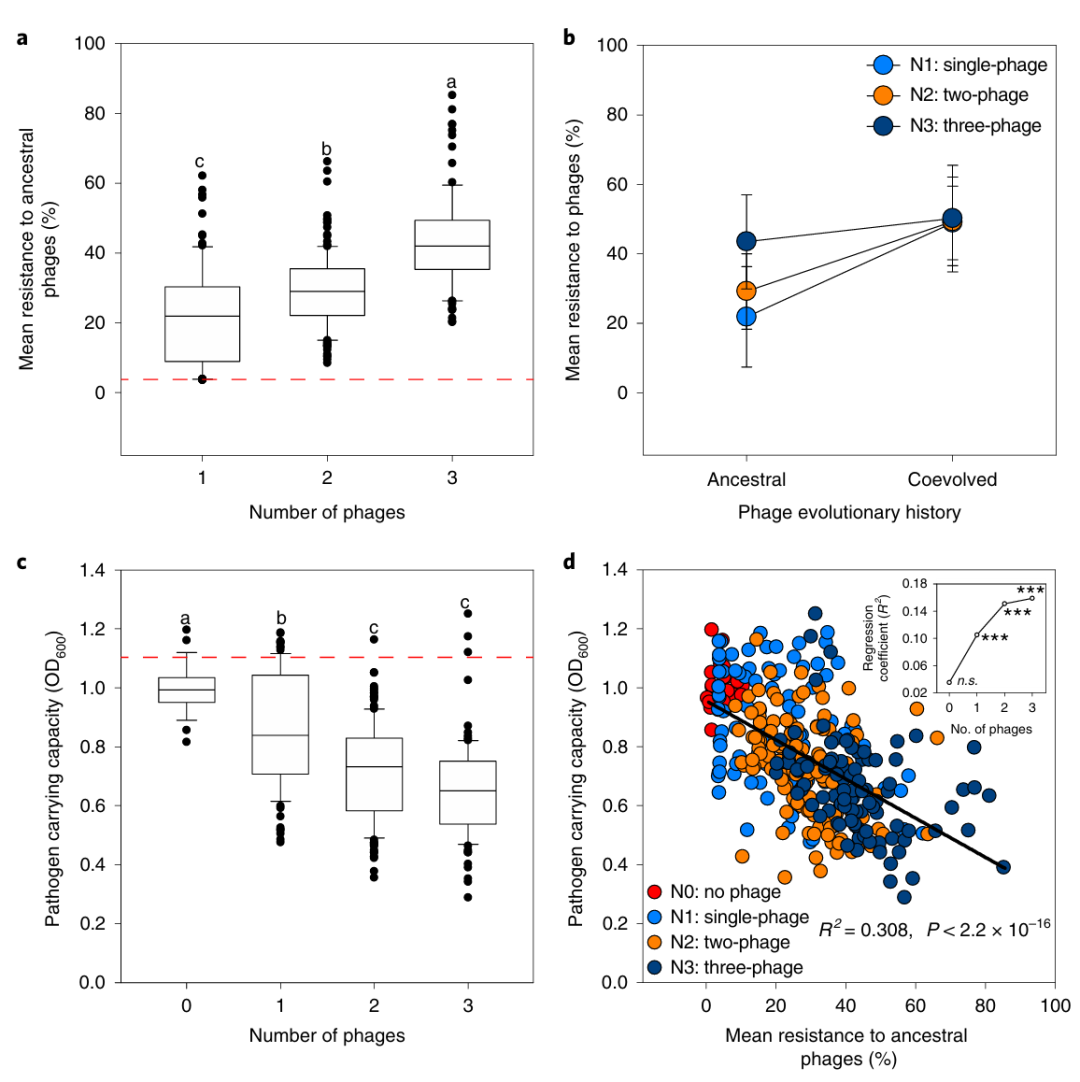
图6 病原菌对原始和共同进化噬菌体的抗性进化(Wang et al., 2019)。(A)病原菌对原始噬菌体的平均抗性(3种噬菌体组合组抗性率最高);(B)为病原菌对共进化噬菌体的平均抗性;(C)为抗性菌株的生长能力(OD600),抗性越高,生长越慢;(D)病原菌生长能力与噬菌体抗性之间的负相关性。
2. 噬菌体与农药协同使用
很多农民习惯只用农药,但研究发现若将噬菌体和农药协同使用,只要搭配得当,效果会远超单独使用。
比如在前面提到关于青枯雷尔氏菌噬菌体LPRS20的研究中,作者发现将LPRS20和噻唑锌铜(常见铜制剂)搭配使用时,如果先喷LPRS20,4小时后再喷噻唑锌铜,对青枯雷尔氏菌的杀灭效果比单独用噬菌体或农药强2倍(图7)。而如果同时喷施农药和噬菌体,农药会暂时抑制噬菌体繁殖,效果反而变差。
另一组搭配是LPRS20和低浓度小檗碱(植物源农药)一起使用,会产生“协同作用”——细菌数量比单独用噬菌体明显减少。但要注意,小檗碱浓度不能太高,否则会抑制噬菌体的繁殖,产生拮抗作用。
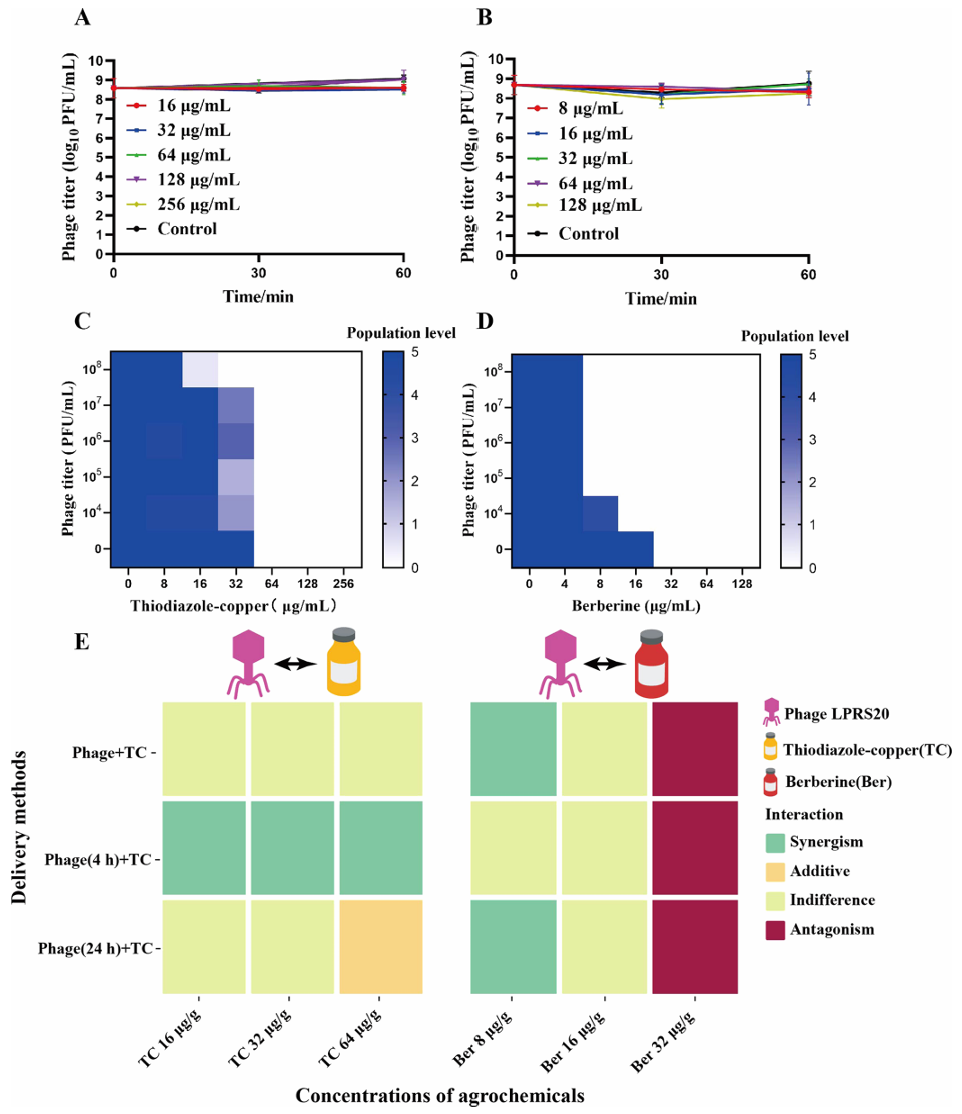
图7 分离的噬菌体LPRS20和噻二唑铜或小檗碱的联合作用(Liu et al., 2024)。(A、B)LPRS20在噻二唑铜(A)和小檗碱(B)中的稳定性;(C、D)LPRS20和噻二唑铜(C)或小檗碱(D)的组合在96孔板的NB培养基中抑制LRS6的生长;(E)噬菌体LPRS20与噻二唑铜或小檗碱的相互作用。
三、噬菌体应用遇到的问题
虽然噬菌体很厉害,但要真正从实验室走向田间应用,仍面临一些挑战。不过在科学家们不断的研究过程中已经找到不少解决办法,让小远带大家详细了解一下:
难题 1:太阳紫外线会“晒死” 噬菌体
噬菌体的DNA很怕紫外线(尤其是UV-B),在叶片上晒2小时,活性就会降低90%。这也是早期噬菌体田间效果差的主要原因。
解决办法:给噬菌体“穿防晒衣”
· 天然提取物:胡萝卜、红辣椒提取物中的类胡萝卜素,能吸收紫外线;酪蛋白、大豆蛋白也能形成保护膜,让噬菌体在叶片上的存活时间从2小时延长到24小时(Halawa, 2023)。
· 高分子材料:海藻酸盐、聚谷氨酸(γ-PGA)这些可降解聚合物,能把噬菌体包成“微胶囊”,既抗紫外线,又能缓慢释放(Haq et al., 2024)。比如用2% γ-PGA包裹噬菌体,在30℃、UV照射下,72小时后活性还能保留60%,而没有包裹的噬菌体只剩10%。
· 避开日晒时间:傍晚或清晨喷施噬菌体,此时紫外线弱,噬菌体有足够时间附着在叶片或土壤中(Buttimer et al., 2017)。
难题 2:在土壤和叶片上存活时间短
根际土壤里的黏土颗粒会吸附噬菌体,叶片上的干燥环境会让噬菌体失活。
解决办法:在喷噬菌体的同时,加入无毒菌株(比如不致病的青枯雷尔氏菌突变体),这些菌株能作为噬菌体的繁殖宿主,让噬菌体在土壤中持续增殖。实验显示,加了无毒菌株后,噬菌体在土壤中的存活时间从7天延长到30天,而且能随着根系生长扩散传播(Haq et al., 2024)。
另外,把噬菌体施到土壤里,比喷在叶片上的存活时间更持久。在之前研究中发现,土壤喷施噬菌体7天后叶片上还能检测到10⁴ PFU/g的噬菌体;而叶面喷施噬菌体,2天后就检测不到噬菌体了。
难题 3:宿主范围太窄,“漏网之鱼” 多
有的噬菌体只侵染一种类型的菌株,遇到其他菌株无法侵染,导致病害复发。
解决办法:把3~4种不同宿主范围的噬菌体混合,覆盖更多菌株。比如针对番茄细菌性斑点病,科学家混合了5种噬菌体,能杀90%以上的病原菌菌株,田间防治效果比单一噬菌体高50%(Buttimer et al., 2017)。
现在还可通过基因测序,分析田间病原菌的基因型,针对性搭配噬菌体,实现“精准打击”(Haq et al., 2024)。
难题 4:储存运输不方便
液态噬菌体在25℃下存放1个月,活性就会下降50%,这给偏远地区的农民带来麻烦。
解决办法:做成“噬菌体粉末”。用冻干技术(加乳糖、海藻糖作为保护剂)把噬菌体变成粉末,在4℃下能存1年,25℃下也能存6个月,活性损失不到10%(Haq et al., 2024)。将冻干的噬菌体粉末,用清水溶解后,对病原菌的抑制效果和新鲜噬菌体几乎一样。
四、噬菌体产品
目前,全球已有多款噬菌体农药商业化,比如美国OmniLytics公司的AgriPhage,能防治番茄细菌性斑点病、辣椒疮痂病,还获得了有机农业认证(OMRI);匈牙利的Erwiphage专门防治苹果火疫病,效果堪比链霉素(streptomycin),且没有抗生素残留问题(Buttimer et al., 2017)。
在国内,之前提到的LPRS20已经进入田间试验阶段,在广东、云南的烟草田,防治效率稳定在40%~50%,比传统铜制剂高15%;噬菌体组合疗法也在江苏番茄基地试点,亩均减少农药使用量30%,土壤重金属含量下降20%。
未来,随着基因工程技术的发展,科学家还会改造噬菌体——比如给它加一个“溶菌酶基因”,让它裂解细菌的效率提升10倍;或者让噬菌体携带“抗病基因”,帮助植物增强免疫力(Haq et al., 2024)。
本期推文中,小远给大家介绍了噬菌体在现代农业中的应用,以及未来发展的潜力。从1924年人类首次用噬菌体防治白菜黑腐病至今,这一“天然杀手”已从实验室走向田间,不仅能精准杀灭植物病原菌、降低80%的植物病害,还能增加土壤中放线菌等有益菌数量、改善土壤微生物结构,对消费者而言意味着更安全无农药残留的蔬菜,对农民来说能减少农药用量、降低成本、避免土壤退化,也是地球可持续农业的关键一环,可减少化学农药对生态的破坏。尽管噬菌体在应用中面临存活条件、运输方式等难题,但小远相信科学家们会突破重重难关。也许未来某天,当你在超市拿起一颗番茄时,它的生长过程中没有大量使用铜制剂、抗生素,而是靠一群看不见的“微型卫士”守护——这就是科技给农业带来的温柔改变,让我们拭目以待吧~
References:
Buttimer C, McAuliffe O, Ross R P, et al. Bacteriophages and bacterial plant diseases[J]. Frontiers in microbiology, 2017, 8: 34.
Halawa E M. Challenges of bacteriophages application in controlling bacterial plant diseases and how to overcome them[J]. Journal of Genetic Engineering and Biotechnology, 2023, 21(1): 98.
Haq I U, Rahim K, Paker N P. Exploring the historical roots, advantages and efficacy of phage therapy in plant diseases management[J]. Plant Science, 2024, 346: 112164.
Liu H, Gu W, Lu Y, et al. Exploration of Phage–Agrochemical Interaction Based on a Novel Potent Phage LPRS20-Targeting Ralstonia solanacearum[J]. Journal of agricultural and food chemistry, 2024, 72(50): 28005-28018.
Wang X, Wei Z, Yang K, et al. Phage combination therapies for bacterial wilt disease in tomato[J]. Nature Biotechnology, 2019, 37(12): 1513-1520.

















 27
27

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









