 食物需求驱动植物库发育
食物需求驱动植物库发育




食物需求作为植物库发育的驱动因素
1. 引言
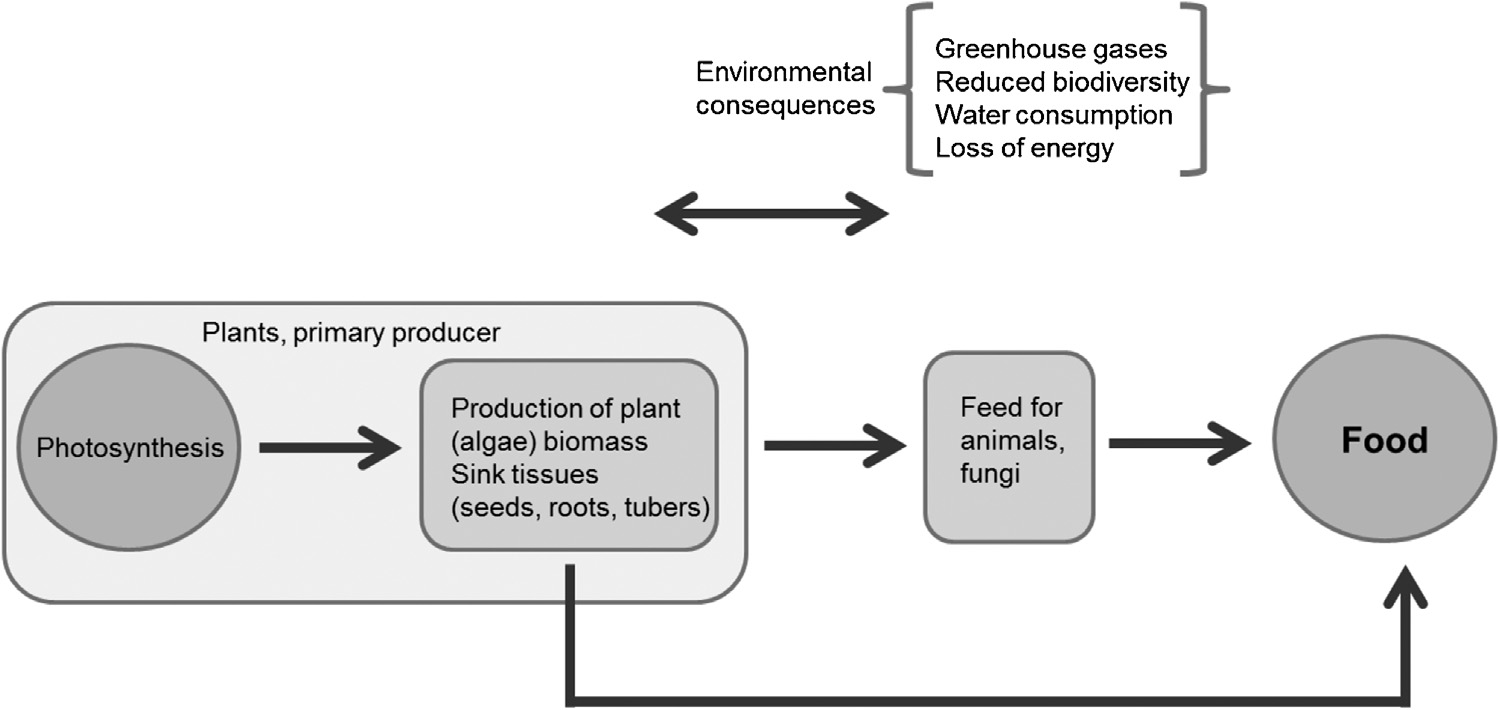
为已经庞大且持续增长的人口进行集约化食物生产对环境造成了负面影响(蒂尔曼等人,2002,2001)。特别是肉类生产大大降低了能源效率,即肉类与植物性产品的消费明显伴随着能量浪费的增加(皮门特尔和皮门特尔,2003)。此外,牲畜甲烷排放增加了温室气体的排放(美国环保局,2010),而为开辟牧场进行的森林开垦则减少了生物多样性,并可能破坏濒危生态系统。
这些负面环境影响只能通过避免过度肉类消费 − 这一社会政治问题 − 和/或在不增加资源消耗的情况下提高每公顷粮食作物产量来缓解(图1)。

2. 食物
被称为“食物”的物质被人类摄入以提供身体所需的营养支持。食物通常来源于植物或动物,少数情况下来源于真菌或细菌。为了饲养动物,必须使用来自动物或植物的饲料喂养它们。因此,从某种意义上说,几乎所有的食物都源自植物,特别是通过一种称为光合作用的过程产生的有机物生产。
相比之下,化学合成过程可能在局部具有重要性,但在全球范围内几乎可以忽略不计(卡伦,2001)。
食物可以直接食用,如黄瓜等蔬菜、苹果等果实、玉米等种子或生菜等叶片。在其他情况下,食物需要经过加工才能成为可食用的产品:(i)通过烹饪,例如马铃薯或水稻;(ii)通过研磨制成面粉,例如小麦、马铃薯或木薯;(iii)通过提取获得如果汁等产品;(iv)通过腌制、加工保存、泡渍、干燥、发酵或烟熏。这些制造技术的发展是为了使食物能够更长期地保存。
源自植物的间接食物消费包括动物产品,如牛奶及其加工制品(例如凝乳或奶酪)、鸟类的蛋以及蜜蜂生产的蜂蜜。除了动物产品外,动物自身的多个部分(统称为肉类)也是食物来源。
3. 库
植物的库器官是光合产物的净输入者(霍,1988),即这些器官无法产生足够的光合物来维持其自身的代谢和生长,必须通过糖类或相关物质形式输入碳。而其他植物器官,主要是成熟叶片,则是光合物的净输出者,即它们产生的光合产物多于自身代谢所需。这些所谓的源叶向库器官供应碳,促进库的生长发育以及库中贮藏物质的积累。植物体内除碳以外的其他物质(如氮或磷)的分配对植物生长也具有重要意义,而对于氮而言,还关系到蛋白质的贮藏(张等,2015;辛克莱和德威特,1976)。
所有地下器官在植物生长期间都是碳库,因为它们显然无法进行光合作用。此外,未成熟叶片以及其他地上器官如花、叶柄、茎或种子也是库。在上述提到的库中,有一些在植物发育过程中会转变为源器官。
叶片成熟后,会经历从库到源器官的转变(特吉恩,1989)。另外,像种子、块茎、根和茎等贮藏库在发育过程中也会由库转化为源。在此过程中,其在库阶段积累的贮藏物质会被再动员并输出,以驱动例如萌发等过程。这种转化通常发生在母株死亡后,或经过一段休眠期后独立于母株发生。一个有趣的例外是热带作物木薯。当木薯遭遇极端干旱条件,即在旱季开始及期间,它会脱落几乎全部叶片以节约水。
当条件再次变得更有利时,即雨季开始时,贮藏根和茎会暂时转变为源器官,以促进新叶片的生成。随着这些新长出的叶片成熟,它们再次成为源器官,而贮藏根和茎则重新转回库状态(El‐Sharkawy, 2004;Hahn 等, 1992)。
4. 食物 − 库 关系
总体而言,植物(以及其他界别的生物体)进化出了两种截然不同的策略以确保下一代“世代”的生存:一种是产生大量个体后代,例如拟南芥所采用的策略;另一种则是仅产生少量后代(Gnan 等, 2014)。这些少数后代通常具备充足的能量储存,有助于其在下一代生长期中建立并存活。在不同物种中,还演化出了介于这两种极端策略之间的多种中间形态。此处的“世代”也可以指像马铃薯块茎这样的克隆,它们与母株在遗传上完全相同,但由母株产生以确保在下一个生长期的存在。
即使植物进化出前一种策略,它们也会为后代(通常是种子)提供贮藏物质,这些物质可在萌发期间被降解,并用于建立能够进行光合作用的幼苗。
此外,植物通过不同的策略实现其后代的传播。在库的背景下,最有趣的一种策略是果实的发育。在这里,种子被包裹在果实中,果实由开花后的子房形成,并充满糖分。这些果实吸引动物食用,随后被排泄出来,从而确保了后代的传播。
早期的人类通过狩猎动物和采集可食用的植物部分来生存。当人类定居下来从事农业时,他们开始驯化动物和植物。为此,他们首先选择想要饲养的动物物种和想要种植的植物物种。随后,他们开始根据自身需求对这些生物体进行优化。
植物中最有营养且可食用的部分通常是库,因为它们含有贮藏物质,可为下一代植物的良好生长提供保障,或吸引动物帮助传播。这些贮藏物质如淀粉、糖、油/脂肪和蛋白质也对人类营养有益,因此驯化(以及后来的育种)最初集中于扩大库,方法是从群体中挑选库最大的个体植株进行进一步培育。
5. 食品改良与植物育种的里程碑
在谷物栽培的早期阶段,通过选择育种塑造了现代作物野生近缘种的基因库,在此过程中,禾本科植物单粒种子和穗的尺寸增加,从而形成了如单粒小麦(Triticum monococcum)等早期谷物。现代小麦(Triticum aestivum)起源于单粒小麦或乌拉尔图小麦(Triticum urartu)与拟斯卑尔脱山羊草(Aegilops speltoides)杂交形成的异源四倍体硬粒小麦(Triticum turgidum),随后该硬粒小麦又与粗山羊草(Aegilops tauschii)杂交,形成异源六倍体普通小麦(Triticum aestivum),并由此开始被广泛种植(国际小麦基因组测序联盟(IWGSC),2014)。从二倍体到 − 在此情况下为 − 六倍体的倍性水平提升,在比较祖先品种与现代品种时常可观察到,并且通常与产量增加相关。马铃薯是另一个比较原始品种与现代品种时表现出倍性水平与产量正相关的例子。然而,倍性水平的提高并非祖先作物产生高产后代的必要条件。全球种植面积和产量最大的粮食作物玉米(Zea mays)为二倍体,其起源于同样为二倍体的大刍草(Doebley, 2004; Beadle, 1980)。这两个谷物例子的共同特征是种子增大以及穗轴脆性降低,从而实现更高效率的收获和脱粒。相反,对于非驯化形态而言,脆弱的穗轴具有优势,因为它有助于更有效的脱落(Li 和 Gill,2006)。
在18世纪中后期工业化时代开始时,通过正向和负向选择进行育种已是农业中的先进技术(Breseghello, 2013)。随着工业化进程的发展,越来越多的人迁入不断扩张的城市,成为工人,因而依赖他人生产其所需食物。此外,工人家庭也在小块园地中而非田地上自产食物。在这两种情况下,都必须提高每公顷产量以养活不断增长的人口。由于谷物产量无法满足需求,一种对欧洲而言较新的作物——马铃薯,开始广泛推广。当时马铃薯相比谷物具有多项优势:更有营养、更易种植、无需研磨,且最重要的是− 其每公顷产量高于谷物,并且至今仍然如此(http://faostat3.fao.org/home/E)。这些优势导致主要社会底层的工人及其家庭完全依赖马铃薯,例如爱尔兰有三分之一的人口依赖马铃薯作为热量来源。在这种背景下,1840年代由卵菌致病疫霉(Phytophthora infestans)引起的马铃薯晚疫病席卷欧洲,造成连续多年的全面收成损失,在欧洲尤其是爱尔兰引发了严重饥荒。爱尔兰人口因此减少了超过20%,数百万人死亡或移民离开爱尔兰(Kinealy, 1994)。
格雷戈尔·孟德尔(1866年)发现“可遗传性状”为有目的的杂交育种——即组合育种——奠定了基础,尽管在相关机制被发现之前,已有自发或人为进行的杂交行为。通过将同一物种的两个品种(或基因型)进行杂交,可获得具有稳定且理想性状组合的第二代后代。
这一方法的改进是培育杂交种,即两个不同品种杂交产生的第一代后代。20世纪初,杂种优势被描述为“特定杂交后代的表现优于双亲”的现象(舒尔,1908年),也就是说,可以培育出比稳定品种产量更高的杂交种。如今,由于杂交种具有更高的产量优势,多种作物已作为杂交种进行种植,其中包括甜菜、油菜和玉米。随着在玉米中开发出母系遗传的细胞质雄性不育(CMS)技术,导致花粉不育,从而不再需要耗时费力的去雄操作来防止自花授粉(施耐布尔和怀斯,1998年)。
杂交玉米的培育及其利用是20世纪60年代作物生产力大幅提高的一个方面,如今这一时期被称为“绿色革命”(埃弗森和戈林,2003年)。另一个重要方面是矮秆谷物的培育及其随后在农业中的推广应用,这些矮秆谷物具有较低的赤霉素(GA)浓度(彭等,1999年)。与它们的祖先相比,这些谷物具有两大主要优势:第一,植物能够将更多的光合产物用于生成可收获器官,而不是投入到长茎的生长中;第二,茎秆的自支撑能力增强,从而在恶劣天气后表现出更高的产量,因为长秆品种在此类条件下容易遭受涝害。此外,随着人工肥料在此期间的开发,作物植株的矿质营养供应也得到增加,其对产量的积极影响(斯图尔特等,2005年)被充分加以利用。
光合产物及其他物质在植物器官之间的分配优化过程尚未完成。相反,源库能力在多种特定环境条件下远未达到优化。
农业集约化的一个重要弊端是对生物多样性的负面影响。(i)为了经济地生产更多食物,需要更多的土地。这些土地以前往往未被用于农业,而是作为避难所(弗霍文和塞特,2010年),(ii)大规模的化学投入能够更好地应用于大面积的单一作物,而不是农业集约化之前农场典型的 小块土地 。(iii)育种者集中于少数几个在早期驯化过程中被选中的作物品种,这些品种特别适合用于集约农业。一些可能具有重要性状(如对病原体的抗性)的源可能已经灭绝(坦克斯利和麦库什,1997年)。
6. 现代食品改良方法
由于全球人口不断增长(联合国,2015)、人均卡路里消耗持续增加(http://faostat3.fao.org/home/E)以及耕地流失问题同时加剧(Pimentel 和 Burgess,2013),提高植物产量的需求日益迫切。从植物生理学家的角度来看,可采用综合策略来弥补这一供应缺口:通过转基因、基因组编辑和育种手段开发产量更高的植物。这些方法应同时推进,即不仅独立应用,还应相互结合,使一种方法中获得的知识和原理能够融入其他方法中。例如,从因某个基因过表达而具有更高产量的转基因植物中获得的知识,可转化为与该基因高表达相关的标记,并用于S.M.A.R.T.(标记辅助与先进繁殖技术)育种,也称为精准育种(Davis 等,1997)。此外,近年来已开发出多种基因组编辑方法,这些方法将在植物育种和生物技术中得到应用并必将被充分利用。在最近的一篇综述中(Cardi,2016),基因组编辑被描述为“一组技术,能够 在特定靶位点对感兴趣的特定基因组序列进行编辑、删除、替换或插入”。
这些新开发方法的一个优势在于,植物基因工程可以以定向方式进行。在此前发明能够利用巨核酸酶(Daboussi 等,2015)、锌指核酸酶(ZFNs;Petolino,2015;Kim 等,1996)、转录激活因子样效应物核酸酶(TALENs;Christian 等,2010;Boch 等,2009)或成簇规律间隔短回文重复序列(CRISPR)相关核酸酶(CRISPR/Cas9; Kumar 和 Jain,2015;Bortesi 和 Fischer,2015;Jinek 等,2012)引入序列特异性DNA双链断裂的方法之前,被概括为“基因组编辑”的技术在高等植物中的应用因同源重组频率极低而难以实现基因靶向。因此,转基因植物(包括突变体)不得不通过 T‐DNA随机插入基因组的方式产生。根据联合研究中心科学与技术报告,大多数基因组编辑技术产生的作物无法与常规育种作物区分开来,因此无法检测(Lusser等,2011)。在这种情况下,有人建议将基因组编辑的植物视为非转基因植物。此外,最终基因组编辑可能提高植物育种的社会接受度(Araki和Ishii,2015;Huang等,2016),尤其是在欧洲。

通过组合式利用这一最新开发的技术组合,迫切需要的食物改良可能比预期更快地实现。
7. 源与库之间的碳分配
如何提高作物产量?要回答这个问题,不妨换个角度思考:高产的主要障碍是什么?对此问题的回答多种多样。举几例而言,(i) 不利环境条件(热胁迫、冷害、涝害或干旱),(ii) 不良农业实践(肥料不足、土壤耕作不当)或 (iii) 各类病害。这些对产量有负面影响的共通之处在于它们都属于环境问题。为了尽量减少这些障碍的影响,从而稳定产量,目前已采取多种措施,例如田间覆盖、人工灌溉、施肥管理、喷洒杀菌剂、除草剂和杀虫剂,或种植抗病原体植物。相比之下,植物内在因素对产量的负面影响较少受到关注,例如植物体内的养分分配。本文重点讨论源组织和库组织之间的碳分配。
在不同的生长阶段,植物为了最终实现最佳产量具有不同的需求(图2)。萌发后不久,植物应形成叶片和根系,以分别确保光合作用和矿物质吸收,从而实现最佳生长(图2A)。在生殖生长期(图2B),植物会填充其库,以为下一代的良好起始做好准备。因此,大部分同化物都用于此目的。当发生病原体侵袭时(图2C),同化物必须用于抵御病原体,从而导致库中同化物的填充无法达到最佳状态。
植物根据其需求进化出了相应的发育和防御程序。然而,作物通常并不生长在这些发育和防御程序所适应的严酷环境中,而是通常受到精心照料,例如通过化学物质保护其免受各种病原体侵害,以获得更好的收获。在这种条件下,更高产量的潜力可能会受到限制性内在发育程序的制约,例如较短的库填充期,或因维持不必要的防御途径所致。克服此类限制的一种方法是同时提高源和库的能力,最终增加碳向库器官的流动,从而获得更高产量的植物。由于在不同的生长阶段,产量可能在源限制和库限制之间转换,因此提高两者的能力可能使植物生长更加协调,最终实现更高产量。成功解除源和库限制的一个例子是具有增强源库能力的三重转基因马铃薯植株(Jonik 等,2012)。葡萄糖‐6‐磷酸/磷酸转运蛋白(GPT),负责将碳骨架导入淀粉体(Kammerer 等,1998),以及腺苷酸转运蛋白(NTT),负责将能量导入淀粉体(Kampfenkel 等,1995),在块茎特异性B33马铃薯蛋白启动子(Rocha‐Sosa 等,1989)的控制下被过量表达。此前已证明,这两种代谢物转运蛋白活性的提高能够增强库强度和块茎淀粉产量,而单独过量表达任一转运蛋白则不能(Zhang 等,2008)。除了提高库能力外,还通过两种不同方法促进叶片蔗糖形成、减少叶片淀粉形成来增强源容量:(i)通过反义方法在叶片中下调ADP‐葡萄糖焦磷酸化酶,或(ii)在叶肉细胞的细胞质中过量表达可溶性焦磷酸酶。所获得的三重转基因马铃薯植株的块茎淀粉产量翻倍(Jonik 等,2012)。
其他可能增强源或库能力的方法已有研究或可预见其前景(图3)。然而,除上述方法外(Jonik 等,2012),目前尚未见同时提高源和库能力的报道。
为了增强源容量,最明显的方法是提高光合作用。一种大规模的前瞻性方法是为所谓的C3植物配备更高效的C4光合作用。C4光合作用的进化在进化过程中独立发生了多次(Sage等人,2011年),并且对从 C3到C4光合作用转变的建模表明,每一个必要步骤都带来了改善,解释了这种进化发生的原因和机制(Heckmann等人,2013年)。雄心勃勃的 “C4水稻”项目旨在将C4光合作用引入最重要的主食之一——水稻,正是基于这一策略(http://c4rice.irri.org/)。其他更为实际的、以改善光合作用为目标的方法也已成功实施。在烟草中过表达拟南芥景天庚酮糖‐1,7‐二磷酸酶(SBPase)和/或果糖‐1,6‐二磷酸酶(FBPase),以及异位表达集胞藻属PCC 6803的无机碳转运蛋白B(ictB),均导致光合性能和生物量增加,在三重转基因植株中尤为显著(Simkin等人,2015年)。在水稻中也进行了类似的方法,其中双功能 SBPase/FBPase和ictB被过表达。此处,光合性能同样得到了提升。然而,在决定水稻籽粒产量的三个参数中,即分蘖数、每穗实粒数和粒重,仅在双重转基因植株中观察到分蘖数增加(龚等人,2015年)。
在另一种方法中,过表达RubisCO活化酶(Rcase)——一种激活 RubisCO的酶——在正常温室条件下没有产生影响。然而,在变化的光照条件下,转基因植物中的RubisCO激活速度更快。此外,过表达一种更耐热的Rcase在轻度热胁迫下实现了更好的RubisCO激活。由于CO2和O2在水中的溶解度随温度升高而降低,导致溶解的CO2与O2比例下降,光呼吸随温度升高而增强。然而,有趣的是,耐热型Rcase的表达似乎有利于光合作用的耐热性(Carmo‐Silva等人,2015年及其参考文献)。为了尝试阻止光呼吸,研究人员在马铃薯(Nölke等人,2014年)和生物燃料作物亚麻荠(Dalal等人,2015年)中过表达了乙醇酸脱氢酶(GlyDH),这是一种由大肠杆菌GlyDH操纵子酶GlcD、E、F组成的嵌合酶。两种情况下的实验结果相似:马铃薯植株表现出块茎产量显著增加,亚麻荠则表现出单株种子重量增加。
尽管大多数提高源容量的方法都集中在酶促反应上,但另一种方法可能是调节代谢物分布,从而改变代谢物依赖性调控。在这方面,人们怀疑胞质糖(尤其是己糖)的水平与光合作用活性呈负相关。为了降低胞质葡萄糖水平,利用强组成型35S‐CaMV启动子在拟南芥tmt1‐2突变体背景下过表达拟南芥的液泡膜单糖转运蛋白1(TMT1)。TMT1将己糖从细胞质转运至液泡,使得这些己糖或其产物不再能被糖感应系统识别。因此,抑制光合作用的信号应会缺失。这种信号变化反映在已知受糖调控的基因的去调控上。已知被糖抑制的叶绿素a/b结合蛋白1(CAB1)在过表达植株中的表达高于对照植物,而糖诱导的硝酸还原酶1(NR1)在TMT1过表达植株中的表达则下调。此外,韧皮部装载蔗糖转运蛋白SUC2在转基因植物中的表达更高。TMT1过表达植株种子产量的增加因此可归因于潜在更高的光合作用以及所产生的蔗糖更有效的韧皮部装载(Wingenter等,2010)。
为了提高库容,除了上述提到的增强与块茎淀粉形成相关的关键代谢物转运蛋白的过表达外,还增强了马铃薯块茎中的蔗糖合酶活性,从而提高了块茎淀粉含量和单株干重(Baroja‐Fernández等,2009)。此外,下调马铃薯质体型腺苷酸激酶(ADK)导致腺苷酸类物质积累,并同时增加了块茎淀粉含量和产量(Regierer等,2002)。另一种方法是通过下调葡聚糖水二激酶(GWD)来减少淀粉储存作物中的淀粉周转。该方法此前在马铃薯中已实施,甚至早于该酶实际活性被发现之前,结果使块茎淀粉含量增加(Lorberth等,1998)。除马铃薯外,最近已在甜菜主根的储存液泡中鉴定出负责蔗糖装载的蛋白质(Jung等,2015)。这种称为TST的蛋白质与前述拟南芥TMT1高度同源,但其转运蔗糖而非葡萄糖。提高TST活性有望增加甜菜及其他糖储存作物的库容。
8. 下一代食物
除了单纯提高食物的可获得性外,作物育种如今更加关注食品质量。这一植物改良的重要进展源于一项发现,即不仅食物的数量,其成分也会影响人类健康。因此,当今发展中国家的主食生产已包含提升食品营养成分的策略。在接下来的几十年中,几乎所有类型的主食都将推出所谓的(生物)强化品种,以应对营养不良相关疾病和早亡问题(http://www.harvestplus.org/)。该方法的先驱是为预防儿童因饮食中缺乏维生素A而导致失明和死亡而研发并推广的“黄金大米”(Ye等,2000)。其他提升食品质量的例子包括富含具有保健作用花青素的番茄植株(Butelli等,2008)以及生物可利用维生素B6含量提高的木薯植株(Li等,2015)。
然而,设计和制定养活全球的策略时,必须考虑目标地区人们所遵循的习惯和信仰。因此,自然科学提供的“简单”解决方案必须得到社会政治项目的支持,以推动教育和启蒙,从而应对当前最紧迫的问题——世界人口过剩。否则,满足更多食物需求的恶性循环只会导致人口进一步增加,进而需要更多的食物。


















 2万+
2万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









