



第8章 纳米技术在细胞内运输、成像和治疗剂递送中的应用
1. 引言
跨细胞膜的药物递送已受到科学家的广泛研究。然而,关于治疗剂在细胞内环境中的命运的研究却相对较少。本综述讨论了纳米技术领域的进展,重点关注药物在细胞内环境中的定位,特别是其在成像和诊断中的作用。可以说,治疗剂对亚细胞器的特异性靶向以及诊断剂已成为药物递送的新前沿。已在纳米尺度范围内研究了多种制药平台,如脂质体、碳纳米管、量子点、纳米胶束和树状大分子,以评估其靶向亚细胞组分的能力。在靶向细胞器的纳米载体领域已取得显著进展。有必要深入了解纳米载体的大小和形状对细胞内运输和分布的影响。同时,确定治疗剂在细胞内环境中是否能够被可控释放也至关重要。如果成功,可将纳米载体导向细胞内细胞器,使治疗剂在该处发挥药理作用。对这些问题及其他类似问题的解答将有助于更好地理解靶向细胞内药物递送。
要回答这些问题,需要实时追踪纳米载体的命运。成像技术的快速发展将使科学家能够探索这一研究领域。本章将回顾的一些令人振奋的技术进步包括最先进的共聚焦显微镜、量子点和金纳米颗粒。这些技术将使我们能够确定纳米载体在细胞内环境中随空间和时间的分布。共聚焦和荧光显微技术的快速进步为科学界提供了一种有效工具,用于成像载药纳米载体进入细胞内环境及在其内的转运过程。延时和Z轴堆叠图像提供了有关细胞内环境以及纳米颗粒和其他纳米载体递送的详细信息。量子点也被称为纳米晶体,由半导体材料制成,逐渐在纳米技术研究中占据了重要地位。关于量子点的详细讨论超出了本章的范围,但将探讨其在细胞内成像和递送中的作用。与量子点类似,金纳米颗粒是另一种重要的成像和递送工具。
药物纳米载体提供了一个理想的平台,可在不改变药物分子本身的情况下改变药物的分布。可以在构成纳米载体系统的组分上进行不同的化学修饰,并将治疗剂载入其中。这将实现靶向治疗,而无需对药物分子本身进行结构修饰。
大多数药物纳米载体都经过表面修饰,从而实现靶向递送。另一种不涉及纳米载体修饰的靶向递送方式是尺寸依赖性靶向。例如长循环脂质体和纳米颗粒等纳米载体可能能够被动靶向具有渗漏血管的组织和细胞,这得益于增强的渗透性和滞留效应。这两种不同的靶向策略可分别分为主动和被动靶向,本章将对此进行讨论。
细胞内部环境与水缓冲溶液大不相同。在缓冲环境中,治疗性分子可以自由扩散并自由相互作用。细胞内环境包含细胞骨架网络和众多细胞器。细胞质中含有大量大分子,如蛋白质和碳水化合物。因此,治疗性分子的扩散或转运在这种黏稠且拥挤的环境中,分子的行为将不同于其在缓冲溶液中的行为。通常认为药物分子在细胞内环境中的扩散是受阻扩散,反映了高度的分子拥挤。
此外,细胞质环境的黏稠性以及药物分子与细胞内组分的结合也可能影响药物在细胞内的扩散。在设计纳米载体时,另一个需要考虑的重要参数是治疗性分子的理化性质。它在决定药物分子的亚细胞命运中起着重要作用。
本章将回顾纳米载体的另一个重要作用,即靶向药物递送至不同细胞器。特别关注细胞核,因为它是许多重要抗癌药物(如阿霉素)的作用位点。如何有效地将纳米载体输送到其在细胞内的靶向作用位点,是本文综述中讨论的一个重要概念。使用不同的靶向部分来特异性地将纳米载体导向细胞器,这些靶向部分包括肽、抗体、脱氧核糖核酸、核糖核酸及其他分子。深入了解这些靶向部分对于理解纳米技术如何有效实现靶向细胞内递送至关重要。
1.1 细胞内运输中的纳米技术
纳米载体(如纳米颗粒、纳米胶束和脂质体)因其能够将治疗剂递送至细胞内区室而被广泛研究。有必要了解这些纳米载体穿过质膜屏障进入亚细胞空间的进入机制。本节详细阐述了这些机制。
2. 机制
2.1 网格蛋白介导的内吞作用
纳米颗粒等货物通常通过网格蛋白介导途径跨质膜(PM)转运 [1]。网格蛋白介导的内吞作用(CME)始于细胞质中的网格蛋白在质膜特定区域聚集[2]。这些网格蛋白被隔离在被称为网格蛋白包被小窝(CCPs)的质膜区域。随后,CCPs发生内陷,形成独立的囊泡,并从质膜脱离,形成网格蛋白包被囊泡。存在一些衔接蛋白,负责传递导致网格蛋白包被载体形成的分子信号。这些衔接蛋白与多种调节蛋白(如AP180和epsin)相互作用,调控网格蛋白包被小窝的形成、囊泡从质膜的脱离以及这些囊泡与细胞骨架的不同相互作用 [3]。
2.2 小窝介导的内吞作用
小窝是质膜上的突起结构,大小范围为50–80 纳米 [4]。它们负责通过非网格蛋白依赖途径摄取纳米载体。结构上,小窝由胆固醇、鞘脂、糖基磷脂酰肌醇锚定蛋白以及整合膜蛋白小窝蛋白‐1和小窝蛋白‐2[5,6]组成。大小为 ~100纳米的颗粒已被观察到利用小窝介导的途径进入细胞内区室。最后,这些纳米载体的摄取通过囊泡从质膜脱离完成,类似于CME [7,8]。
2.3 网格蛋白和小窝蛋白非依赖性机制
不依赖网格蛋白和小窝的途径是当前研究的活跃领域。关于哪些途径依赖或不依赖网格蛋白和小窝,目前尚无共识。然而,大多数科学家认为,总体上可分为三种途径。普遍认可的不依赖网格蛋白、小窝和巨胞饮作用的途径包括 RhoA介导的摄取和非网格蛋白依赖载体。
2.4 RhoA介导的摄取
该途径由GTP酶RhoA调控。该途径最初是在研究白细胞介素‐2受体b链( IL‐2Rb)的内化过程中发现的。该途径独立于网格蛋白,因为IL‐2Rb的摄取受到Rac1、p21(RAC1)激活激酶1(PAK1)和PAK2[10]的影响,而这些因子不参与网格蛋白介导的内吞作用。
2.5 非网格蛋白依赖载体/糖基磷脂酰肌醇锚定蛋白质富集的早期内体区室途径
另一种非网格蛋白和非小窝依赖的机制是CLIC/GEEC通路。从结构上看,CLIC/GEEC通路由一条 w40 nm宽的管状突起以及GTP酶调节因子组成。它与焦点黏附激酶‐1(GRAF1)和磷脂酰肌醇‐4,5‐二磷酸3‐激酶相关[11]。这些膜载体受到cdc42和ARF1的抑制。
2.6 浮舰蛋白介导的内吞作用
浮舰蛋白是嵌入质膜中与小窝蛋白‐1相似的蛋白质。这些蛋白质在质膜内呈现为有组织的结构域簇。据观察,由flotillin组成的结构域以网格蛋白非依赖的方式从质膜上脱离。囊泡进入细胞质环境后,flotillin存在于多泡结构内部,这些结构与内体过程密切相关[12,13]。
有研究表明,浮舰蛋白介导的内吞作用在纳米颗粒摄取过程中起着关键作用。多项研究显示,浮舰蛋白参与了多聚复合物和二氧化硅纳米颗粒的摄取。这些研究为浮舰蛋白在负责纳米颗粒进入和转运的内吞作用过程中的作用提供了证据 [14,15]。
3. 巨胞饮作用
经典的巨胞饮作用是一种不依赖于网格蛋白和小窝的机制。它通过巨胞饮体的形成,摄取大量液体和细胞外物质[16,17]。在结构上,巨胞饮作用由膜褶皱的形成以及经典液相标记物(如辣根过氧化物酶和葡聚糖)的摄取所构成[18]。该摄取机制由一组小 GTP酶(如Ras、cdc42和Rac)介导[19,20]。巨胞饮作用可自发发生,也可由激活受体酪氨酸激酶的生长因子诱导而发生的影响触发。这些GTP酶和激酶负责信号转导诱导肌动蛋白调节的途径 [21]。这反过来会诱导巨胞饮杯的形成。据报道,一些阳离子纳米颗粒通过小胞饮作用被摄取。纳米载体的表面电荷影响其摄取机制[22]。
3.1 吞噬作用
吞噬作用的机制与巨胞饮作用类似,都涉及肌动蛋白的聚合,从而引起细胞膜皱褶。当树突状细胞和巨噬细胞等吞噬细胞 engulf 大于 0.5mm[23]的颗粒时,该过程发生。金和银等各种纳米颗粒通过此途径进入细胞内环境 [23]。
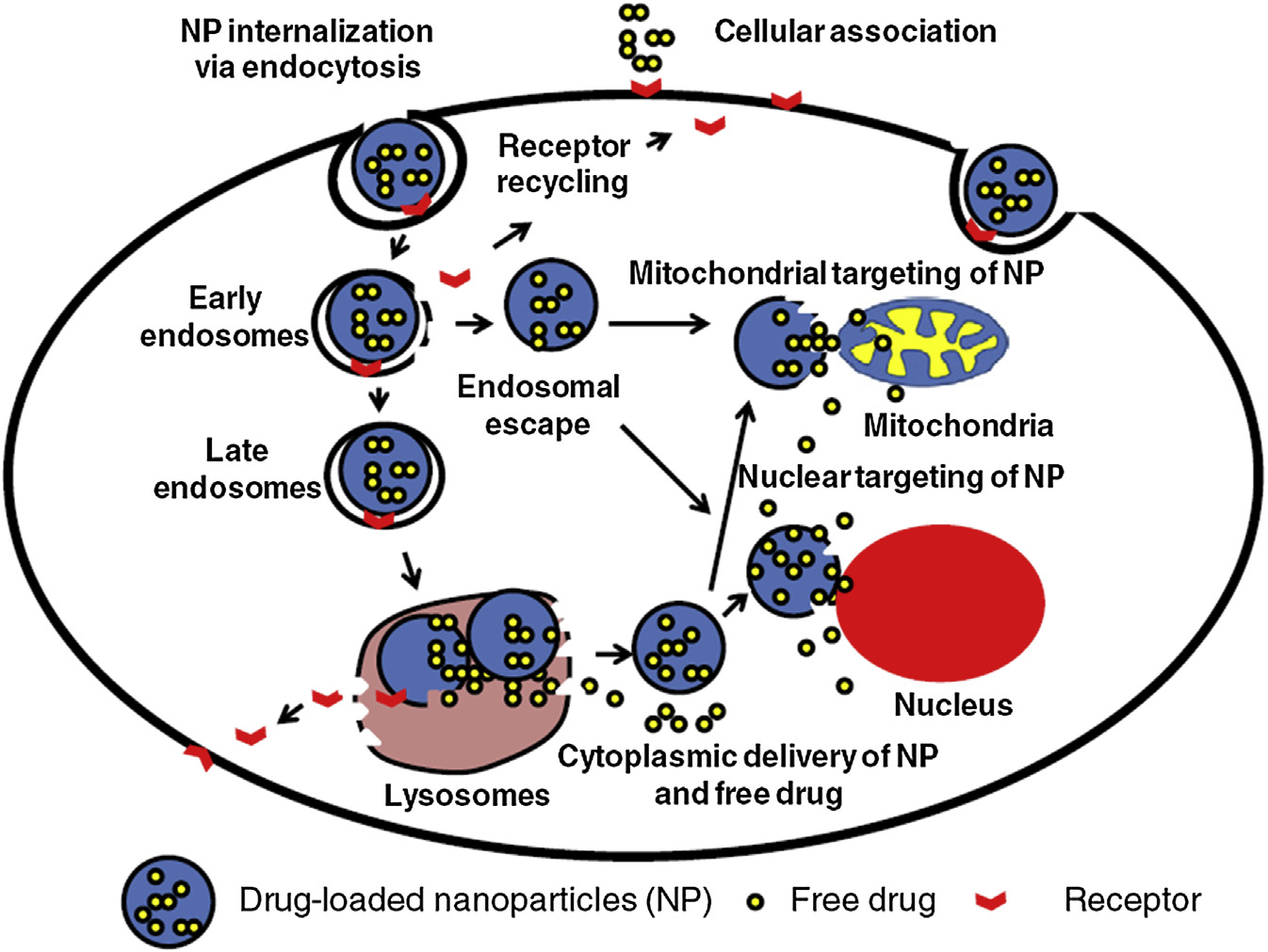
3.2 纳米材料细胞内运输的不同阶段
如前一节所述,纳米颗粒利用多种内吞途径进入细胞内区室。摄取后,纳米颗粒会沿着特定的内吞途径进行转运。纳米颗粒进入细胞内区室的机制是一个关于内吞囊泡内蛋白质含量的动态过程。在摄取过程中,多种衔接蛋白以及效应蛋白会从含纳米载体的囊泡结构表面发生共轭或交换[24]。内吞囊泡的融合及囊泡成熟过程可能会改变含纳米颗粒的囊泡上的蛋白质含量性质。此过程始于细胞内运输的早期阶段。早期内体和巨胞饮体融合形成新的囊泡[25]。
3.3 早期内吞囊泡
初始阶段的内吞囊泡是细胞内运输早期阶段结构多样的群体。早期内体由特定的标志蛋白(如EEA1或Rab5)表征,并以不同的亚群形式共存[26,27]。据观察,成熟速度较慢的内体其运动性降低;相反,快速成熟的囊泡则具有高度的运动性[28]。
3.4 晚期内体‐多泡体
晚期内体也被称为多泡体(MVBs)[29]。在大多数情况下,这些囊泡由早期内体形成。这些是生理上重要的事件,可能调节许多重要过程,如细胞信号传导或胞吐作用。结构上,MVBs是直径为400–1000纳米的细胞质细胞器,由平均大小约为50纳米的内体腔内囊泡(ILVs)组成[30]。随着MVBs与质膜融合,内体腔内囊泡以 Rab27b蛋白 调控的过程作为外泌体释放出来 influenced byRab27b protein[31,32]。已观察到缺乏NPC1的细胞比健康细胞在内体系统中保留纳米载体的时间更长[33]。该结果表明NPC1依赖性的胆固醇稳态在纳米颗粒的细胞内转运途径中具有重要作用 pathways of nanoparticles [34]。
早期和晚期内体在细胞内药物递送策略中均发挥重要作用[35]。靶向线粒体和细胞核的药物利用此途径[36]。
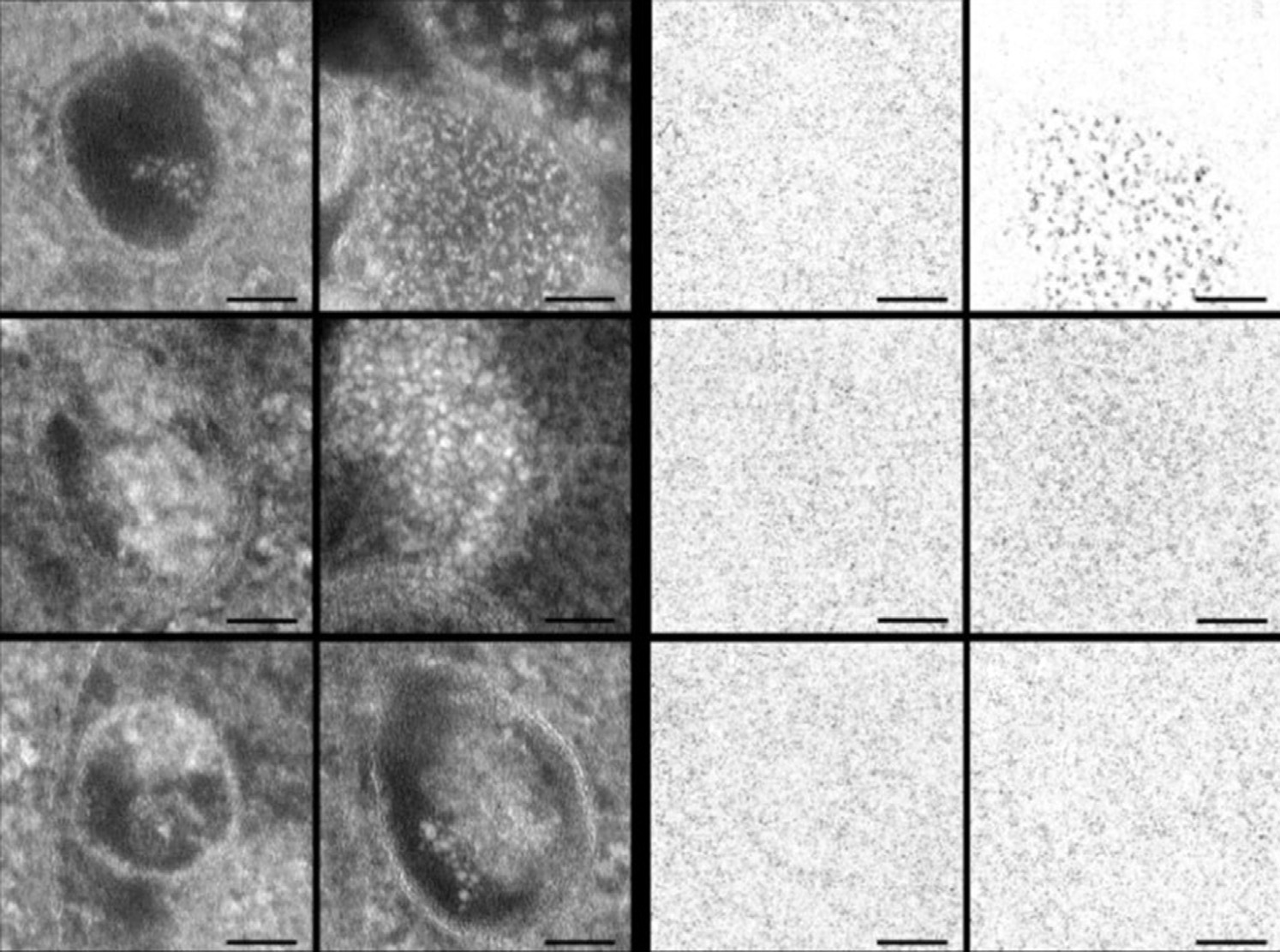
3.5 溶酶体
溶酶体被认为是内吞作用以及自噬途径中的最终细胞器。这些囊泡是多种组分的最终归宿。溶酶体可影响诸如表面受体表达和抑制微生物感染等生化事件[39,40]。显微镜检查表明,溶酶体的结构和体积是非均一的。溶酶体内部环境呈酸性,pH值为4.6–5[41]。这种酸性由多种H⁺ ATP酶产生并维持。当细胞经封装甲氨蝶呤(MTX)的pH敏感型壳聚糖纳米颗粒处理后,溶酶体膜会被降解。
 该过程已通过吖啶橙(AO)重定位技术进行研究[42]。在设计纳米制剂时,研究人员需谨慎确保药物递送系统的靶点不是溶酶体。这些颗粒需要在溶酶体形成之前完成内涵体逃逸。一旦纳米结构被溶酶体吞噬,颗粒将被降解,若靶点为细胞核、线粒体或其他细胞器,则无法递送其负载的货物。
该过程已通过吖啶橙(AO)重定位技术进行研究[42]。在设计纳米制剂时,研究人员需谨慎确保药物递送系统的靶点不是溶酶体。这些颗粒需要在溶酶体形成之前完成内涵体逃逸。一旦纳米结构被溶酶体吞噬,颗粒将被降解,若靶点为细胞核、线粒体或其他细胞器,则无法递送其负载的货物。
3.6 纳米材料的细胞内转运
利用多聚复合物的研究揭示了这些纳米结构在内溶酶体囊泡内的主动运动[14]。已有多种方法被采用以将载药纳米载体递送至亚细胞器。由于纳米载体具有颗粒特性,通常会经历内吞细胞进入机制及随后的内溶酶体加工[43]。人们假设纳米结构非常适合将治疗剂递送至内溶酶体系统。目前正针对纳米药物被内溶酶体系统的摄取开展大量研究。该内溶酶体系统可作为载药纳米颗粒的靶点,因为许多疾病与内体和溶酶体相关。它还可能作为将治疗剂递送至线粒体或细胞核等胞内作用位点的载体。通过网格蛋白依赖性受体介导内吞作用进入细胞的载药纳米结构易受到溶酶体降解。通过网格蛋白非依赖性内吞作用摄取的纳米结构可能导致内体蓄积[45]。因此,在构建药物递送平台时,纳米结构的设计必须考虑这一方面。与纳米结构偶联的配体性质会影响药物递送系统是否能成功将其货物递送至内体,或最终在溶酶体中被降解[46]。
细胞质中形成的生物分子通常包含短肽链 [47]。这些短肽序列也被称为“分子邮政编码。”这些分子影响生物分子进入亚细胞器的特异性摄取 [48,49]。这类短肽通常由特定细胞器中的输入机制识别。这些分子邮政编码可用于将纳米结构递送至不同的亚细胞区室。线粒体和细胞核是目前研究中最常被靶向的两种细胞器。原因是这两种细胞器可能对多种疾病的病理学至关重要。因此,这些细胞器是载药纳米结构的理想靶点 [50]。许多经表面修饰并共轭了线粒体趋向性配体的脂质体制剂已提高了抗癌药物的疗效。金纳米颗粒已被广泛用于通过配体(如腺病毒核定位信号(NLS)以及与整联蛋白结合的结构域片段)将其递送至细胞核 [40]。为了增强治疗性DNA进入细胞核的摄取,研究人员构建了一种带有核定位信号肽(PKKKRKVEDPYC)的 capped 3.3‐千碱基对巨细胞病毒荧光素酶‐NLS基因 [51]。由于该肽链的存在,转染效率提高了上千倍。纳米结构可共轭上能引导其趋向靶向细胞器的配体,或制备成对特定细胞器具有固有倾向的颗粒。靶向配体方法的一些实例此前已有提及。第二种方法的一个例子是使用线粒体趋向性两亲性去甲基氯化铵 [52–54]。这种分子可自发组装形成称为DQAsomes的囊泡或基于脱甲基氯喹(DQA)的类脂质体囊泡。DQAsomes由线粒体趋向性分子组成,被认为是用于线粒体靶向递送治疗性大分子的理想载体。这些囊泡也可作为小分子药物递送的优良载体,特别是抗癌治疗剂,这类药物通过在线粒体中发挥药理作用诱导细胞凋亡。另一种线粒体靶向纳米制剂被称为线粒体靶向脂质体[55]。这些囊泡可与线粒体膜融合,并将其内容物释放到线粒体内部环境。因此,这些结构能够将所携带的货物特异性地递送至线粒体。
纳米结构的形状和表面结构在载药纳米结构的亚细胞分布中也起着至关重要的作用 [56]。表面结构的改变可能导致纳米结构对特定亚细胞器趋向性的变化。例如,fluorescein isothiocyanate-标记的层状双氢氧化物(LDH)纳米粒子可被制备成六角片层,其平均直径范围为50–150 nm,厚度为10–20 nm[57]。这些纳米颗粒也可形成直径为30–60 nm、长度为100–200 nm的纳米棒。纳米棒优先沉积于细胞核中,而六角片层则滞留在细胞质中。据推测,微管控制的转运现象导致了纳米棒在细胞核中的快速摄取。

3.7 颗粒进入细胞核的机制
细胞核在药物递送方面可能是最重要的细胞器。它是许多关键抗癌药物(如阿霉素)发挥药理作用的作用位点[58]。它是基因表达的最终作用位点。质粒 DNA通过脂质复合物在内体逃逸后递送的精确机制尚不明确。控制核质间运输的核孔复合体(NPC)在人类和其他真核细胞中起着关键作用[59]。NPC影响通过核膜的主动和被动运输。被动运输是一种非特异性机制,用于运输直径不超过10纳米且分子量不超过25千道尔顿的颗粒。被动扩散不消耗任何能量,因为它从不逆浓度梯度进行。而促进性或主动运输则涉及能量的消耗。在主动核运输中,该能量以鸟苷三磷酸(GTP)的形式提供。主动运输可以顺或逆浓度梯度进行。主动运输可逆浓度梯度工作,其颗粒截断尺寸约为50纳米,分子量范围为25–75千道尔顿。塔琴科等人[61]已预测出适合纳米颗粒靶向核递送的理想截断尺寸。这项研究表明,颗粒理想上应具有小于100纳米的直径以穿过细胞膜,并小于30纳米以通过核孔复合体进入细胞核。
通过核孔复合体的主动运输通常需要将纳米颗粒或质粒DNA与核定位信号偶联。该肽可介导颗粒通过核孔复合体的特异性摄取。该肽能与核膜输入蛋白(如输入蛋白α和β)[62]形成偶联物。另一种类似的肽称为细胞穿透肽(CPP),可使颗粒穿过细胞膜并逃逸内体囊泡[63]。如果载药颗粒或质粒 DNA无法逃逸内体囊泡,则这些结构很可能在溶酶体中被降解。因此,在设计靶向细胞核的载药纳米制剂时,应考虑使用CPP和NLS。
3.8 纳米结构细胞内转运的成像技术
3.8.1 共聚焦显微镜技术
共聚焦显微镜技术,最常见的是共聚焦激光扫描显微镜,是一种能够提高光学分辨率和图像对比度的强大技术。它在物镜的共焦平面上增加了一个空间针孔,这一机制可消除失焦的光线。因此,三维重建成为可能。通过光学切片在不同深度捕获的一系列图像可以被重建为三维图像,从而提供更丰富的结构信息 [64]。共聚焦显微镜技术的进步拓展了对纳米构建物细胞内运输研究的视野。
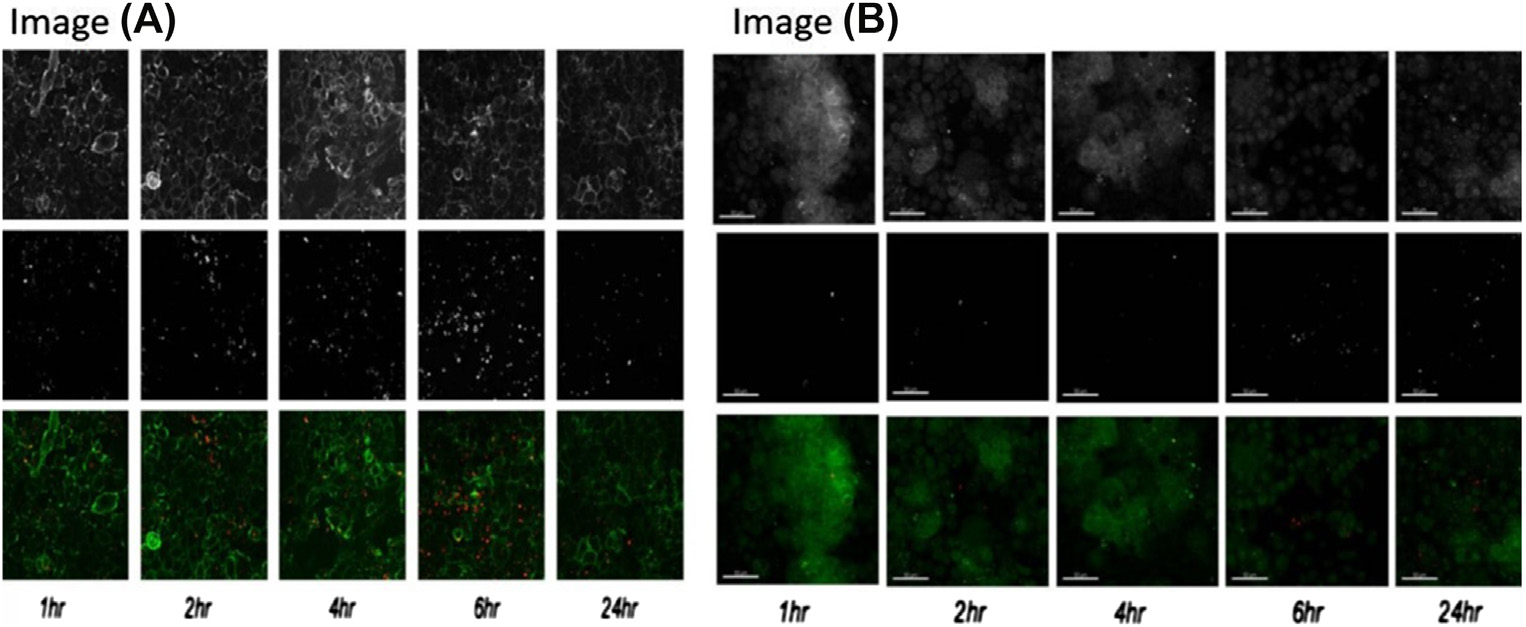
为了更好地评估这一现象,共聚焦显微镜采用了一种称为Z轴堆叠(也称为焦点堆叠)的策略。这是一种数字图像处理方法,将多个在不同焦距下捕获的图像进行组合。该过程生成的合成图像比其组成图像中的任何一个都具有更大的景深。共聚焦激光扫描显微镜结合Z轴堆叠是追踪纳米结构细胞内定位的有力工具 [65]。聚乳酸‐羟基乙酸共聚物(PLGA)纳米颗粒包裹的荧光罗丹明被研究用于在亚细胞器中的分布,借助Z轴堆叠方法的共聚焦显微镜技术进行观察。该成像技术可提供PLGA纳米颗粒在人支气管上皮细胞的高尔基体和早期内体中的定位图像,在溶酶体中的分布则较少[66]。
3.8.2 量子点和金纳米颗粒
量子点是一种微晶半导体纳米结构 [67]。它能够在全部三个空间方向上限制导带电子、价带空穴或激子的运动。已广泛应用于追踪和成像纳米颗粒的细胞内命运。与传统荧光技术相比,量子点的一个显著优势是其不会发生显著的光漂白,这使得研究人员可以自由地设计实验 [68]。然而,量子点确实具有荧光性质(尽管没有显著的光漂白),这使其成为一种有吸引力的诊断和治疗工具。
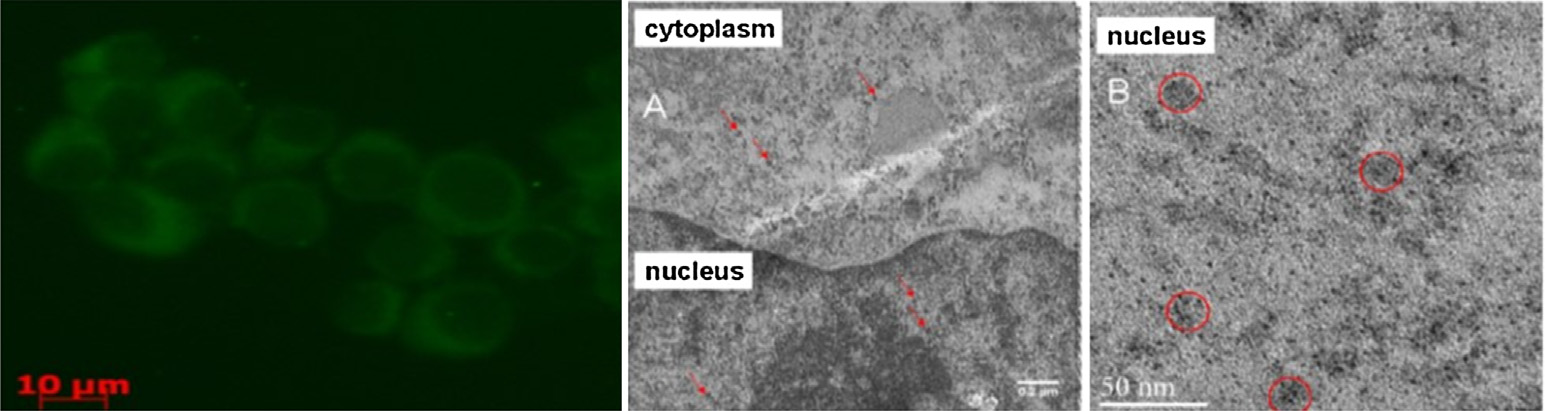
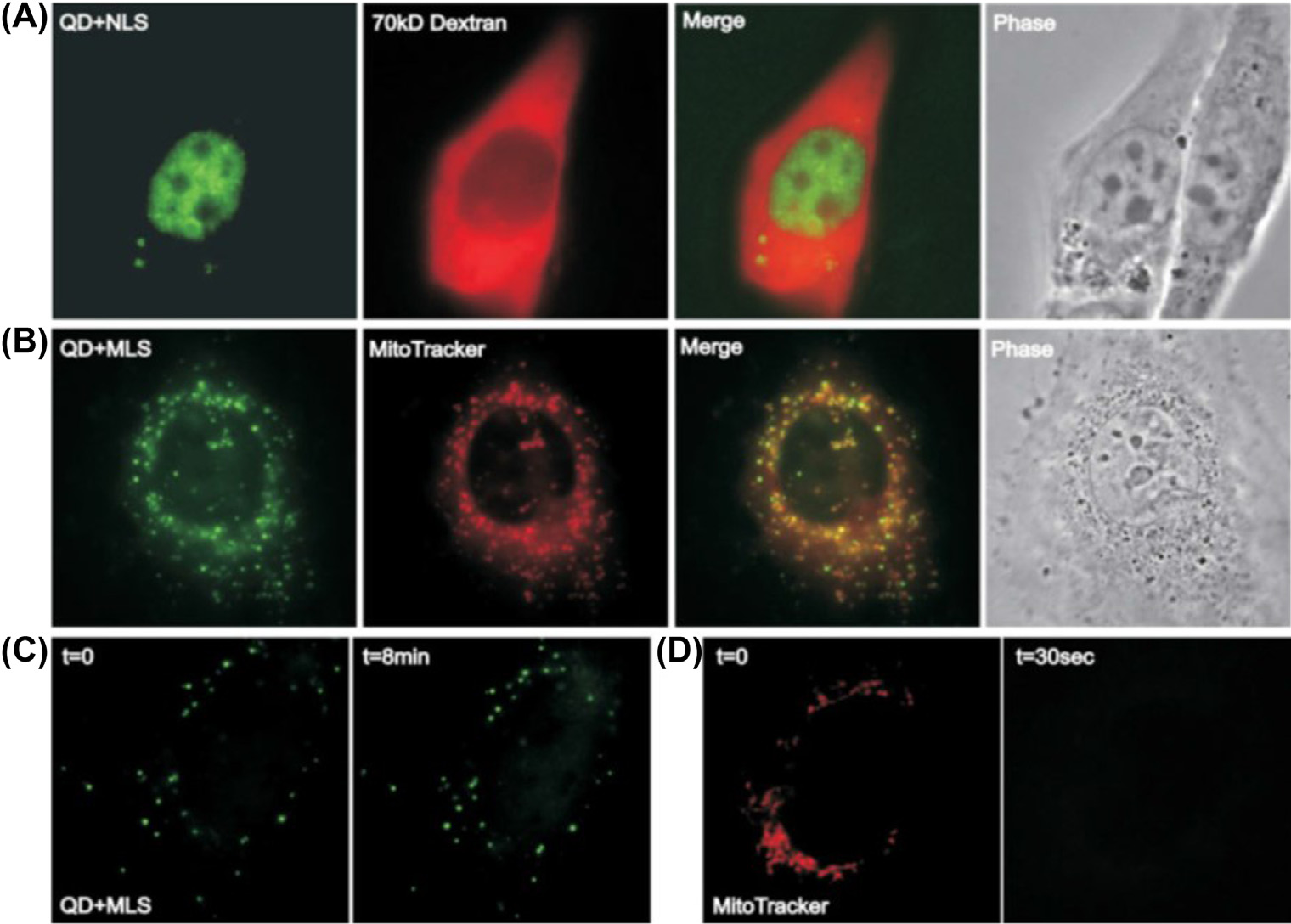
在一项研究中,观察到量子点位于内体和溶酶体囊泡中(图8.8)。这种运输方式模拟了载药纳米颗粒所经过的途径。量子点结合透射电子显微镜技术和共聚焦显微镜技术正在被探索作为研究纳米结构细胞内运输的有力工具。除了内体和溶酶体区室外,量子点还可靶向其他细胞器,如线粒体和细胞核[70]。23个氨基酸的细胞核定位信号和28个氨基酸的线粒体定位信号的共轭可分别导致量子点在细胞核和线粒体中的定位。在细胞摄取开始后30分钟内,量子点即定位于相应的区室[70]。将信号肽与量子点共轭可揭示特定细胞器摄取过程的相关机制和途径。该过程使我们能够开发可特异性靶向特定细胞器的载药纳米结构。肽偶联量子点还使我们能够确定纳米结构在不同靶向细胞器中的定位动力学。
3.8.3 金纳米颗粒
金纳米颗粒被认为是药物和生物医学应用的重要平台。这归因于其理化性质。惰性核心不会产生细胞毒性。金纳米颗粒在可见光和近红外光谱范围内吸收和散射共振光[71]。这些波段的等离子体共振可在广阔的光谱范围内进行优化,方法是通过调节颗粒组分、大小和形状等参数实现 [72]。金纳米颗粒的光散射特性很强,易于检测,其信号强度高于大多数荧光团,并且像量子点一样不会发生光漂白[73]。对于30 nm以下的纳米颗粒,光吸收强于散射。这一现象可用于光热显微镜成像。此外,也可通过透射和扫描电子显微镜轻松成像。因此,金纳米颗粒更适合用于研究颗粒通过核孔复合体的运动机制和动力学[73]。
金纳米颗粒的潜在应用包括但不限于生物成像、单分子追踪、生物传感、药物递送、转染和诊断。将相关配体通过生物偶联连接到金纳米颗粒上,可实现对肿瘤细胞的特异性靶向,同时最大限度地减少对正常细胞的摄取。这有望为肿瘤性疾病提供有效的诊断和治疗策略。 表8.1 展示了金纳米颗粒在与不同配体共轭时的多种亚细胞定位情况。
4. 结论
本综述概述了细胞内药物递送领域的研究进展,并阐述了设计作用位点在细胞内的纳米结构时需要考虑的一些关键参数。如前所述,这一令人兴奋的研究领域仍处于早期阶段,许多重要问题仍有待进一步研究解决。

















 45
45

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









