



传统 ADC 偶联如同撒网捕鱼,疗效难以把控且易出现问题,而定点技术恰似精准制导的导弹!本文将揭开 mTGase、分选酶 A、糖基偶联这三大“魔法师”的神秘面纱,看看它们如何施展定位之术,让药物分子在抗体上有序排列,既稳定精准又长效持久!
传统的 ADC 偶联方法面临着半衰期短、疗效降低以及脱靶效应等诸多挑战。定点偶联技术 (如酶促偶联、糖基偶联等) 提供了精准的解决之道。
在已披露的 576 款 ADC 中,采用定点偶联技术的有 445 款 (占比 77%),占据绝对优势;非定点偶联的仅有 131 款 (占比 23%)。在定点偶联 ADC 管线中,61% 处于活跃开发状态,而非定点管线中仅有 32% 保持活跃。由此可见,定点偶联技术正在加速渗透 ADC 管线。
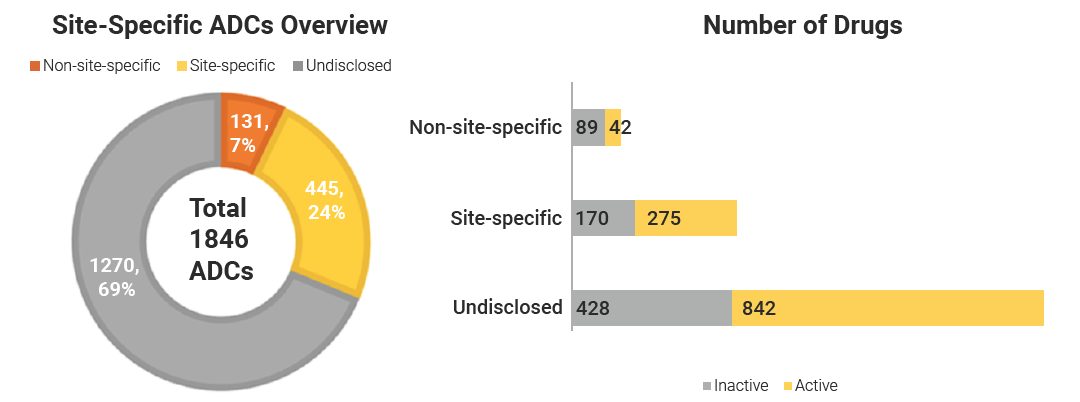
图 1. 定点偶联与随机偶联管线情况。
Section.01
随机偶联 VS 定点偶联
在 ADC 的设计中,诸多因素会对其成功率产生影响,例如抗体、连接子、有效载荷的选择、药物连接位点、偶联方式以及药物抗体比 (DAR) 等。其中,药物与抗体的偶联方式尤为关键,药物的连接位置和方式会对 ADC 的稳定性、疗效等产生影响。
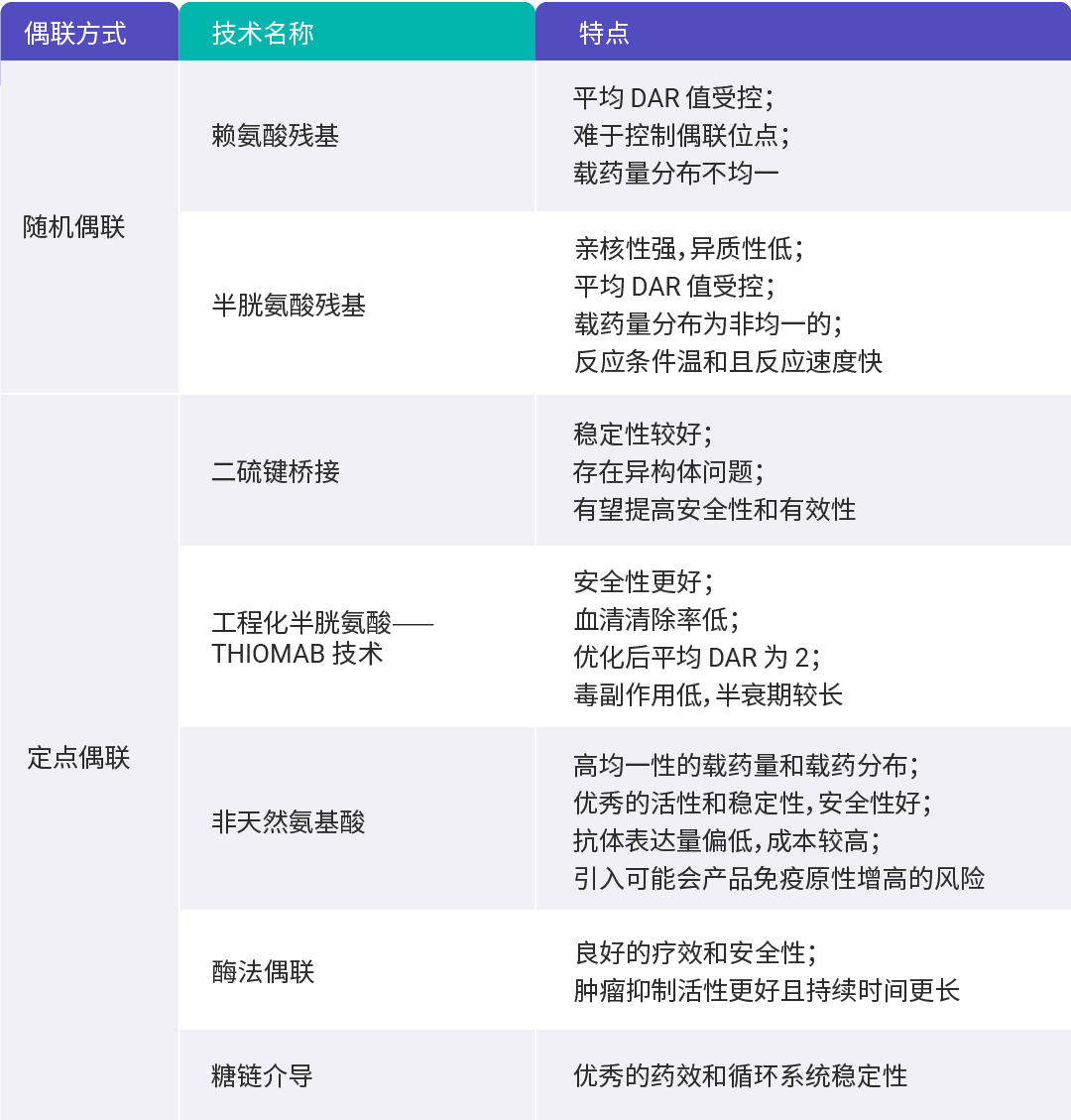
偶联方式主要分为两类,即随机偶联和定点偶联。
随机偶联无需对抗体进行改造,是基于赖氨酸和半胱氨酸的偶联技术,其过程具有随机性,偶联位置和载药数目不确定,所产生的 ADC 是 DAR 不一的混合物,稳定性较差,易发生聚集,动力学性质也不尽相同,从而导致非治疗性毒副作用增加。
而定点偶联技术包括引入反应性半胱氨酸偶联 (Thio - mab) 技术、链间二硫键改造、非天然氨基酸偶联技术、酶法偶联、糖基偶联等。与随机偶联相比,定点偶联得到的 DAR 较为明确,最终成品的一致性更高。
表 1. 不同偶联方式的特点。
Section.02
mTGase 酶促偶联路径
微生物转谷氨酰胺酶(Microbial Transglutaminase, mTGase)能够通过特异性催化脱糖抗体第 295 位谷氨酰胺侧链与含伯胺基团的毒素/连接子形成稳定的肽键,从而实现定点偶联。
mTGase 可以特异性识别去糖基化 IgG 抗体 Fc 区域的 Q295 位点,并将其作为修饰位点与适配底物相结合。通常先利用 PNGase F 进行去糖基化处理,然后再与带氨基的底物发生反应,以此实现偶联[1]。
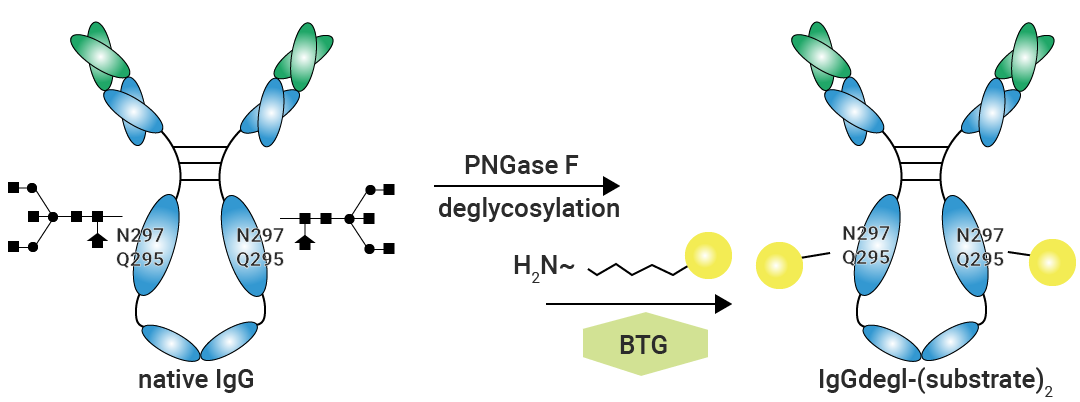
图 2. mTGase 酶促偶联路径[1]。
Section.03
Sortase A 酶促偶联路径
Sortase A (SrtA)是一类由革兰氏阳性菌 (如金黄色葡萄球菌、链球菌等) 分泌的转肽酶。这类酶能够识别特定的氨基酸序列,并催化其与细胞壁或细胞膜形成共价连接。
来自金黄色葡萄球菌的 SrtA 能够特异性识别 C 端含有保守氨基酸序列 Leu - Pro - Xxx - Thr - Gly (LPXTG)、N 端含有多聚甘氨酸的蛋白质 (X 为除半胱氨酸、色氨酸以外的任意氨基酸)[2]。
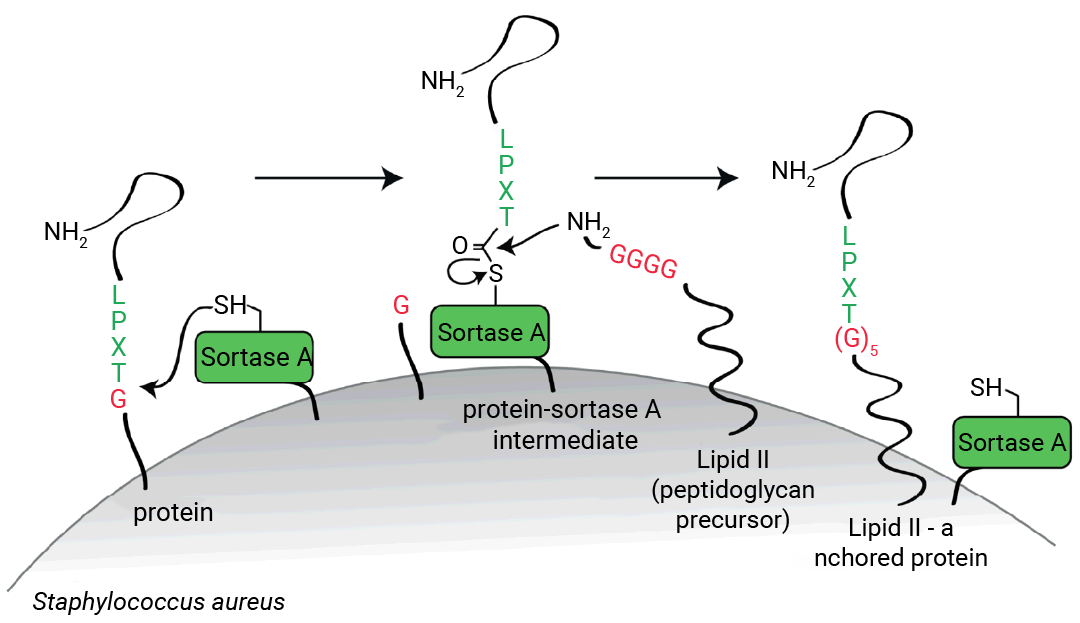
图 3. Sortase A 酶促偶联路径[2]。
Section.04
糖基偶联路径
糖基位点特异性抗体 - 药物偶联物 (gsADCs) 借助 IgG Fc 上的 Asn297 N - 糖基化位点作为药物负载的偶联位点,通常需要多种酶参与糖工程改造[3]。
常见路径为:先运用糖苷内切酶 (如endo s、endo s2、endo SH 等) 对抗体 N - 297 位的糖进行酶切,再利用糖基转移酶 (如GalT(Y289L)) 引入叠氮基,最后通过叠氮与环辛炔反应形成稳定的 ADC。糖基偶联的详细内容可参考往期。
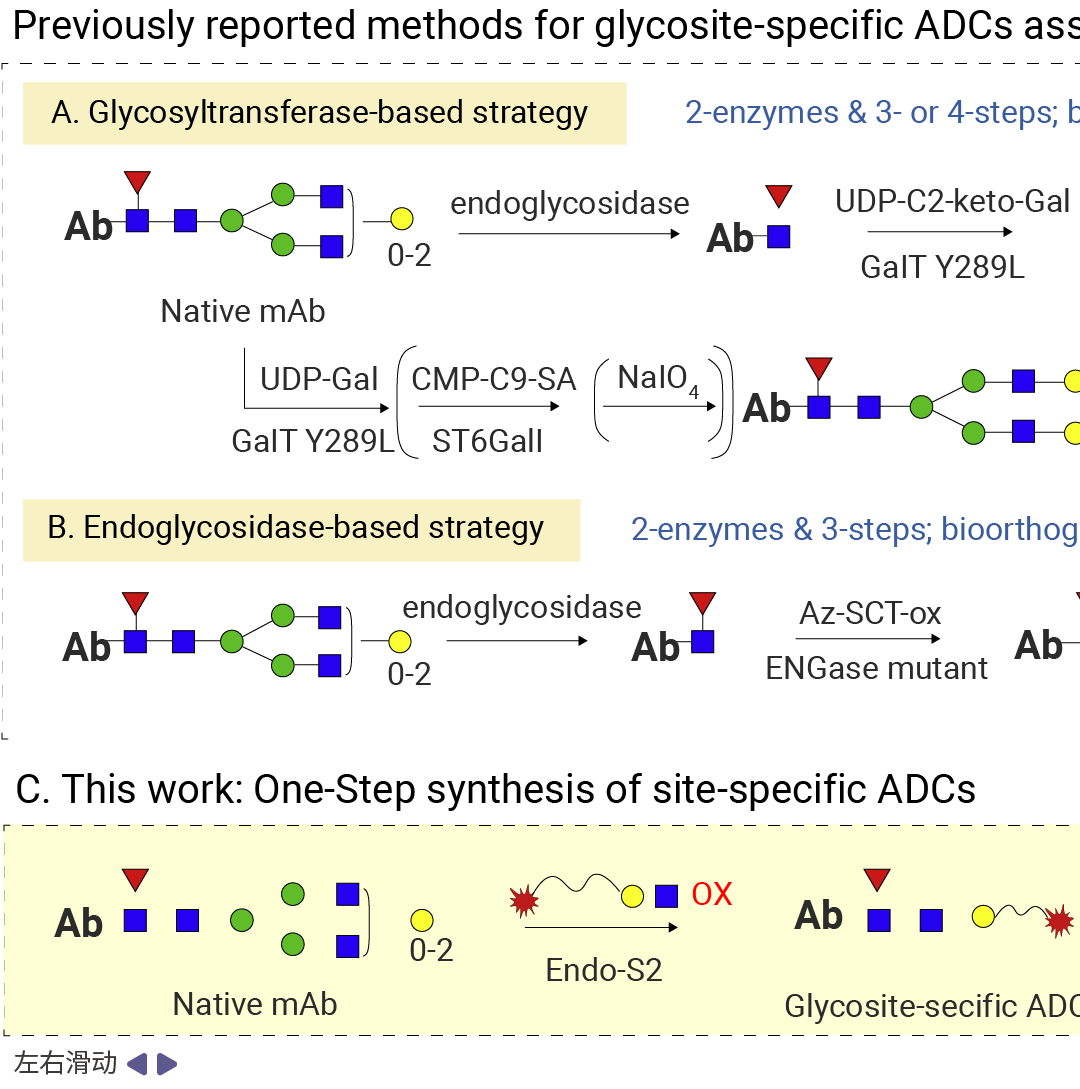
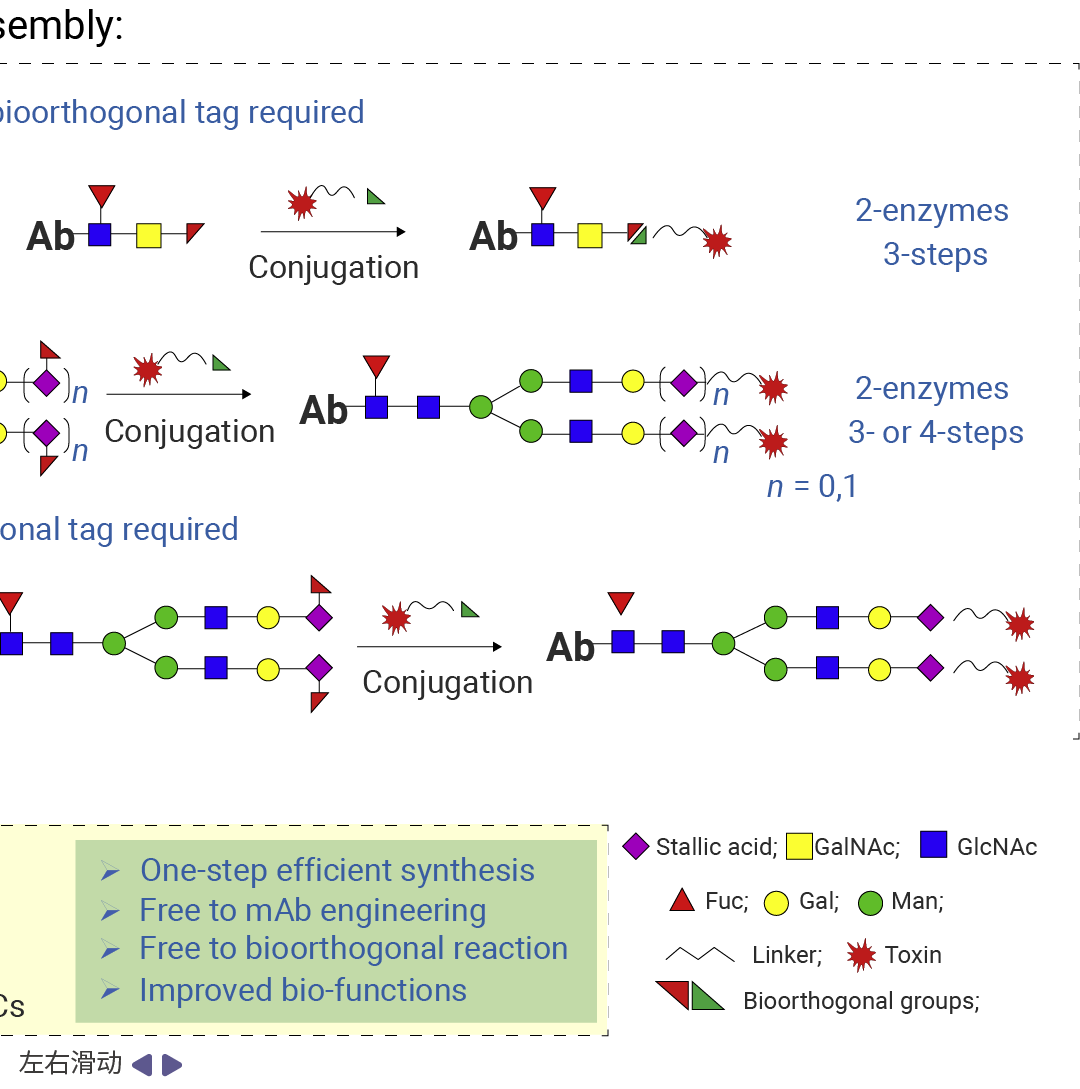
图 4. 糖基偶联路径[3]。
Section.05
小结
小 M 与大家一同学习了糖基偶联与酶法偶联的技术路径。若您认为这些内容有所帮助,不妨点击收藏以备不时之需,更欢迎点赞支持,并分享给有需要的伙伴。您的互动是我们持续输出优质内容的动力,让我们在技术探索的道路上携手共进!
|
产品推荐 |
|
Microbial Transglutaminase Microbial Transglutaminase (MTG) 转谷氨酰胺酶,具有高催化效率、高度的专一性、作用条件温和等特点。 |
|
PNGase F PNGase F 可从糖蛋白中去除几乎所有的 N-连接寡糖。 |
|
PNGase F (Lyophilized) PNGase F 可从糖蛋白中去除几乎所有的 N-连接寡糖。冻干粉形式提供。 |
|
Sortase A, S. aureus Sortase A, S. aureus 在细胞表面各种分子的连接中发挥重要作用。 |
|
Endo S2 一种参与胞质溶胶中游离寡糖加工的关键酶,可催化N-连接寡糖水解。 |
|
Endo S 水解 IgG 抗体 γ 链 Asn-297 上天冬酰胺连接复合型聚糖的 β-1,4-二-N-乙酰壳二糖核心。 |
|
Endo SH 是一种 Endo S 和 Endo H 的融合蛋白,可对 N-糖蛋白中的高甘露糖,复杂型和某些杂合型寡聚糖的壳二糖核心结构进行切割,去除糖蛋白中的 N-连接高甘露糖,可完全替代 Endo S 和 Endo H。 |
|
Bovin beta-1,4-galactosyltransferase 1 (Y289L) 是一种牛来源的在 Y289L 基因位点上发生突变的突变型半乳糖基转移酶。 |

[1] Simone Jeger ,Site-Specific and Stoichiometric Modification of Antibodies by Bacterial Transglutaminase[J].Angew. Chem. Int. Ed. 2010, 49, 9995–9997.
[2] Carla P. Guimaraes, Site-specific C-terminal internal loop labeling of proteins using sortase-mediated reactions[J].Nat Protoc. 2013 September ; 8(9): 1787–1799.
[3] Wei Shia, Wanzhen Lid, Jianxin Zhangd. One-step synthesis of site-specific antibodyedrug conjugates by reprograming IgG glycoengineering with LacNAc-based substrates. Acta Pharmaceutica Sinica B.2022:12(5):2417-2428

















 852
852

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









