






目录
巨噬细胞来源的含 NAMPT 的 EV 介导 NAM 代谢与 CAF 的串扰
巨噬细胞与 CAFs 之间的 NAM 代谢串扰调节 CD8 T 细胞细胞毒性+
含 NAMPT 的 EV 通过 NICD 的 SIRT1 依赖性脱乙酰化抑制 CAF 中 NNMT 的表达
含 NAMPT 的 EV 重塑 TIME 并增强 GC 中的抗 PD-1 反应
摘要
免疫检查点阻断导致晚期胃癌的治疗取得突破。然而,胃癌的突出异质性,特别是肿瘤微环境的异质性,突出了抗肿瘤反应是多因素相互作用的反映的观点。通过转录组学分析和动态血浆样本分析,我们确定了肿瘤微环境中的代谢“对峙”机制,烟酰胺代谢的双重预后意义表明了这一点。具体来说,分别表达限速酶烟酰胺磷酸核糖转移酶和烟酰胺 N-甲基转移酶的巨噬细胞和成纤维细胞调节烟酰胺/1-甲基烟酰胺比率和 CD8 T 细胞功能。从机制上讲,烟酰胺 N-甲基转移酶被 NOTCH 通路转录因子 RBP-J 转录激活,并通过 SIRT1/NICD 轴被含有烟酰胺磷酸核糖转移酶的巨噬细胞衍生的细胞外囊泡进一步抑制。通过自体注射细胞外囊泡纵烟酰胺代谢恢复了胃癌中 CD8 T 细胞的细胞毒性和抗 PD-1 反应。
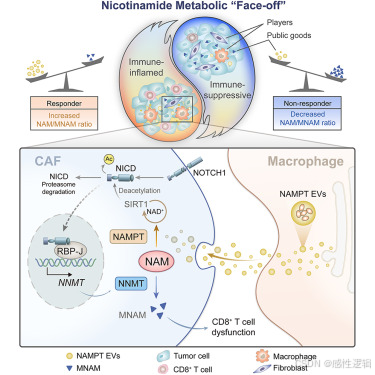
结果
NAM 代谢在 GC 免疫治疗中的双重意义
代谢物是参与代谢串扰的核心中间体。然而,缺乏对代谢物与肿瘤免疫微环境 (TIME) 之间关系的系统评估。在这里,我们根据亚洲癌症研究组 (ACRG 队列,n = 299) 和 PRJEB25780 (n = 45) 数据集的 GC 队列中的代谢物-蛋白质相互作用网络对整体代谢物活性进行了评分。分析了 1,800 多种代谢物和 3,000 种代谢反应的预后意义。
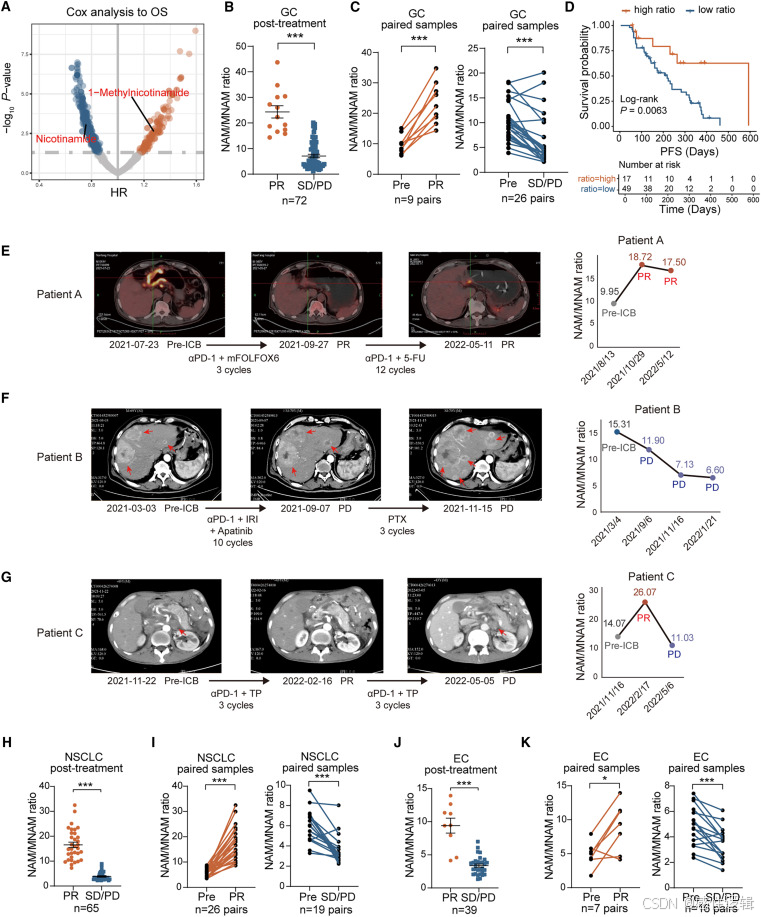
巨噬细胞和成纤维细胞在 GC 中主导 NAM 代谢
代谢物是表征代谢反应活性的下游效应物。在这里,我们专注于参与 NAM 代谢的反应。作为共享底物,NAM 可以由 NAM N-甲基转移酶 (NNMT) 催化,也可以通过 NAM 磷酸核糖转移酶 (NAMPT) 进入 NAD 挽救途径(图 2A)
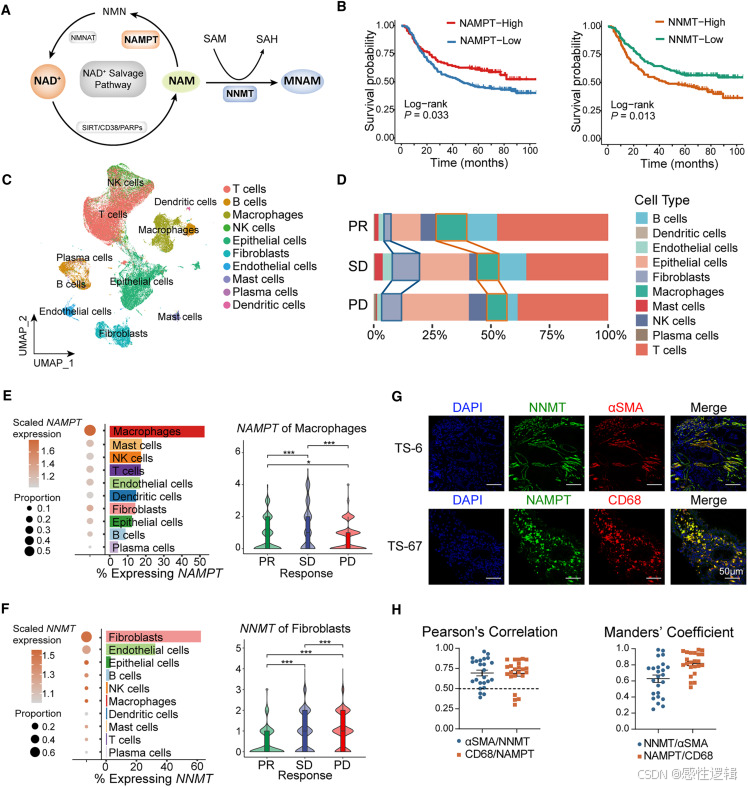
巨噬细胞来源的含 NAMPT 的 EV 介导 NAM 代谢与 CAF 的串扰
研究了巨噬细胞和成纤维细胞通过 NAM 代谢的作用。NAD 代谢,其中 NAMPT 是限速酶,指导巨噬细胞炎症表型的维持和促炎细胞因子的产生。
巨噬细胞与 CAFs 之间的 NAM 代谢串扰调节 CD8 T 细胞细胞毒性+
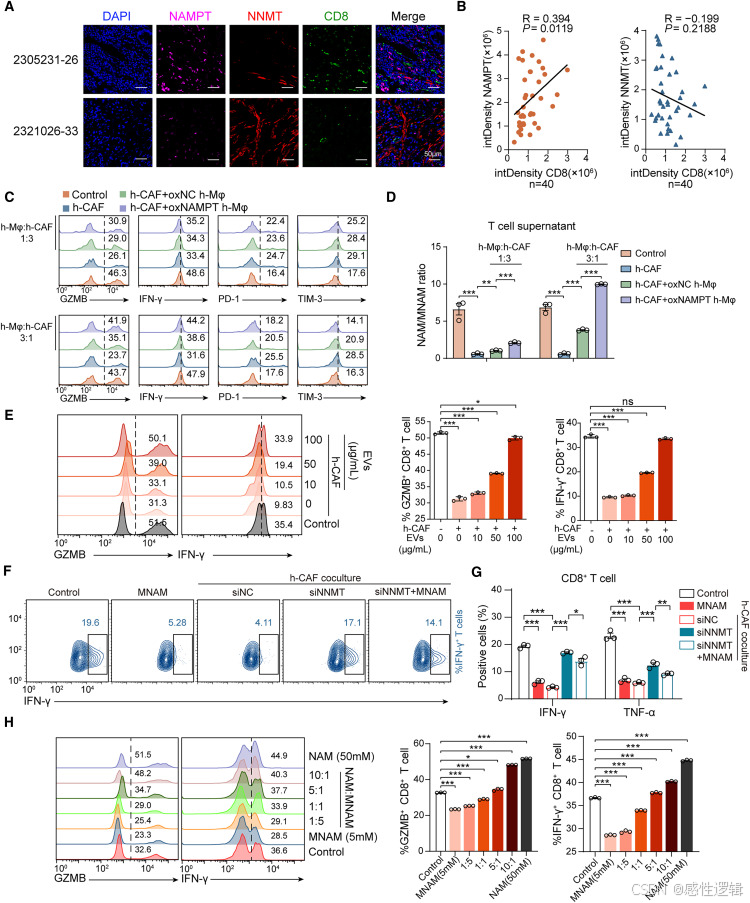
含 NAMPT 的 EV 通过 NICD 的 SIRT1 依赖性脱乙酰化抑制 CAF 中 NNMT 的表达
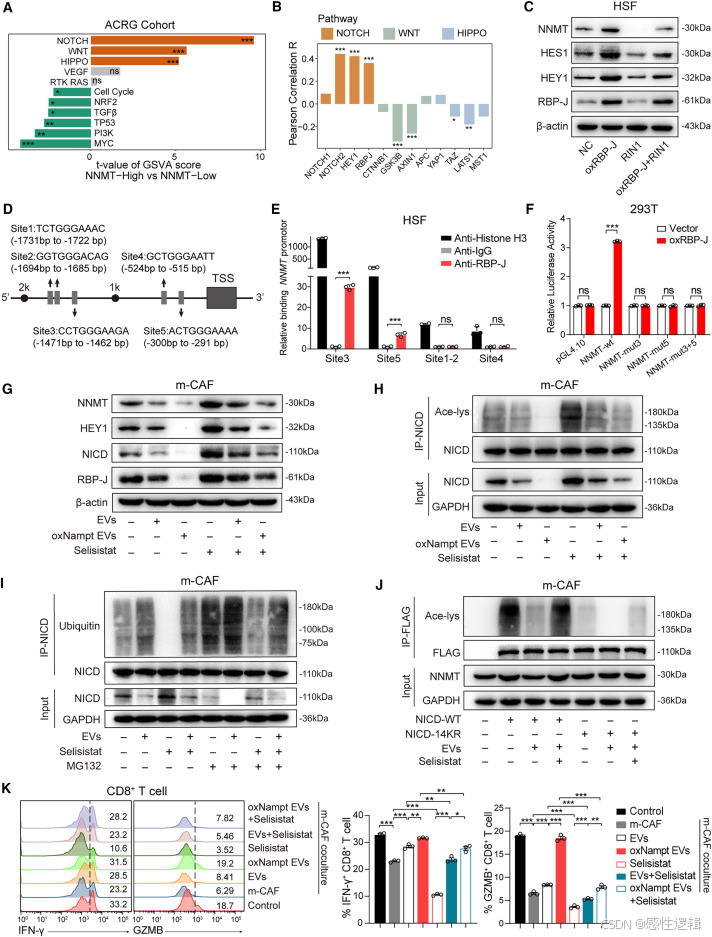
接下来,我们探讨了巨噬细胞来源的 EVs 抑制 NNMT 表达的机制。对 11 种致癌途径的差异分析显示,GC 中 NNMT 高表达组的 NOTCH、WNT 和 HIPPO 通路显著上调(图 5A),其中 NOTCH 通路激活的基因与 NNMT 表达的相关性强于 WNT 和 HIPPO 通路(图 5B)。此外,WNT 激动剂和 HIPPO 抑制剂均不影响 NNMT 表达(图 S6A 和 S6B)。因此,我们认为 NOTCH 通路可能与 NNMT 表达有关。
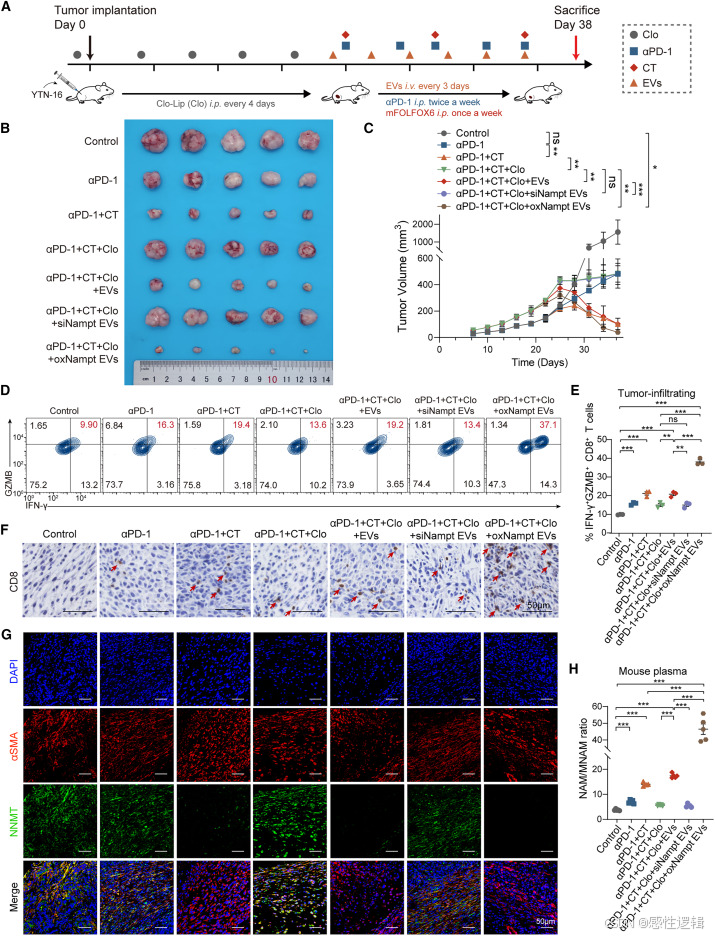
含 NAMPT 的 EV 重塑 TIME 并增强 GC 中的抗 PD-1 反应
参考文献:


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









