







目录
摘要:脑血流(CBF)为脑活动提供了一种定量且可靠的测量手段,并越来越多地被用于研究功能网络。然而,当前的方法主要基于CBF的时间动态来评估区域间的关系,这种方法受到信噪比低和时间分辨率差的限制。在本研究中,我们提出了一种通过估计区域静态CBF概率分布的形状相似性(以Jensen-Shannon散度为指标)来构建功能脑网络的方法,这些静态CBF是通过动脉自旋标记灌注成像在扫描期间测量得到的。基于30名健康受试者在10次访问中提供的CBF数据,我们发现CBF网络表现出非平凡的拓扑特征(例如,小世界属性、模块化结构和枢纽),并且具有低到中等的重测信度和高被试间一致性。我们进一步发现,区域间CBF的相似性依赖于解剖距离,并且在高阶和低阶亚网络之间存在差异。此外,高阶亚网络内的区域间CBF相似性显著低于低阶亚网络。最后,我们发现CBF网络的节点度与区域大小和CBF水平相关,并且与多巴胺转运体和代谢型谷氨酸受体5强度图、主要富集在胆固醇相关通路和内皮细胞中的基因表达水平图,以与记忆、语言和执行功能相关的元分析激活图在空间上一致。总的来说,我们提出的方法为研究人脑功能网络组织提供了一种新颖、相对可靠且具有神经生物学意义的手段。
关键词:脑网络;脑血流量;动脉自旋标记;基因表达;神经递质
1 引言
功能连接已成为表征内在功能动态的有效方法(Van Dijk等,2010),并被应用于评估多种神经精神疾病的改变,例如抑郁症(Ye等,2015;Zhang等,2011)和痴呆症(Pievani等,2011;Wang等,2013),以及与不同认知能力(Li等,2017;Sripada等,2021)和智力(Langer等,2012)的关联,还有在发育(Gao等,2015;Hwang等,2013)和衰老(Sala-Llonch等,2014)过程中的变化。目前,功能连接主要基于功能性磁共振成像(fMRI)测量的血氧水平依赖(BOLD)信号进行估计。然而,BOLD信号是神经活动的间接指标,它测量的是受脑血流(CBF)、血容量和血液氧合影响的血流动力学变化(Hillman,2014)。相比之下,通过动脉自旋标记(ASL)灌注成像测量的CBF是脑功能的直接且定量的生物学指标(Wang等,2008;Wong,2014)。此外,CBF与脑代谢率(葡萄糖和氧消耗)密切相关(Vaishnavi等,2010a)。因此,近年来CBF越来越多地被用于评估功能连接。
迄今为止,已有多种基于CBF估计功能连接的方法被提出。例如,早期研究将独立成分分析应用于CBF时间序列,并识别出5个与BOLD功能网络密切匹配的静息态网络,这些网络在空间和时间特征上高度一致(De Luca等,2006)。随后,Liang等人通过计算区域CBF时间序列的皮尔逊相关性构建了功能网络,并发现其枢纽区域与基于BOLD的功能网络的枢纽区域高度重叠(Liang等,2014a)。基于皮尔逊相关的方法还被扩展到多重回归(Li等,2020)和交叉相关(Zou等,2009)。尽管这些方法取得了成功,但需要注意的是,基于CBF时间序列的功能连接方法受到CBF时间序列信噪比低和时间分辨率差的限制(Alsop等,2015;Wong,2014;Zou等,2009)。这些缺点极大地限制了基于CBF时间序列的功能连接方法的应用场景。因此,有必要开发新的方法来克服这些缺点,以利用CBF研究功能连接或网络。
在本研究中,我们提出了一种通过估计区域CBF概率分布的形状相似性来构建功能脑网络的新方法,这些CBF是基于扫描期间的静态平均灌注图测量得到的。以往研究表明,静态平均灌注图具有高信噪比,并表现出高重测信度(Zou等,2015)。具体而言,我们采用核密度估计方法拟合区域CBF的概率分布,并基于Jensen-Shannon散度(JSD)量化其形状相似性。对于得到的CBF网络,我们系统地表征了其拓扑结构,评估了其重测信度、被试间一致性以及潜在影响因素,并探索了其化学结构、遗传和认知相关性。图1展示了整体数据处理流程。在本研究中,我们假设区域间静态CBF概率分布的形状相似性能够提供一种重测可靠、具有神经生物学意义且与认知相关的研究功能脑网络的方法。
2 材料与方法
2.1 伦理声明
本研究中使用的所有数据集均来自公共数据库。对于每个数据集,受试者的招募过程和知情同意书均获得了相应机构审查委员会的批准。本研究获得了华南师范大学脑研究所与康复中心机构审查委员会的批准。
2.2 受试者与数据采集
本研究使用了一个公开的重测信度(TRT)数据集(Chen等,2015)。该数据集包含30名受试者(15名女性;平均年龄24岁,标准差为2.41),这些受试者均无神经系统疾病或精神疾病史、物质滥用史或导致意识丧失的头部损伤史。每位受试者在一个月内接受了10次扫描,每次扫描间隔3天。具体而言,动脉自旋标记(ASL)灌注磁共振成像(MRI)数据是使用3.0T Discovery MR750扫描仪(美国通用电气公司,威斯康星州,密尔沃基)采集的,采用3D伪连续ASL技术,参数如下:重复时间(TR)=4.834秒;回波时间(TE)=11.088毫秒;翻转角(FA)=111°;层厚=3毫米;无间隔;层数=100;视野(FOV)=相位FOV的100%;平面矩阵=128×128;控制-标记对数=44;总采集时间=411秒;标记持续时间=1500毫秒;标记后延迟时间=1525毫秒。由于这些数据的采集时间早于Alsop等人(Alsop等,2015)推荐指南的发布,因此这些参数并未遵循该指南的建议。在数据采集过程中使用了背景抑制和血管压榨技术。此外,还对每位受试者进行了T1加权快速梯度回波扫描,以获取高分辨率的解剖图像,参数如下:TR=8.1毫秒;TE=3.1毫秒;反转时间=450毫秒;FA=8°;FOV=256×256毫米²;平面矩阵=256×256;体素大小=1.0×1.0×1.0毫米³;176个矢状切片。在数据采集前,每位受试者均签署了书面知情同意书。
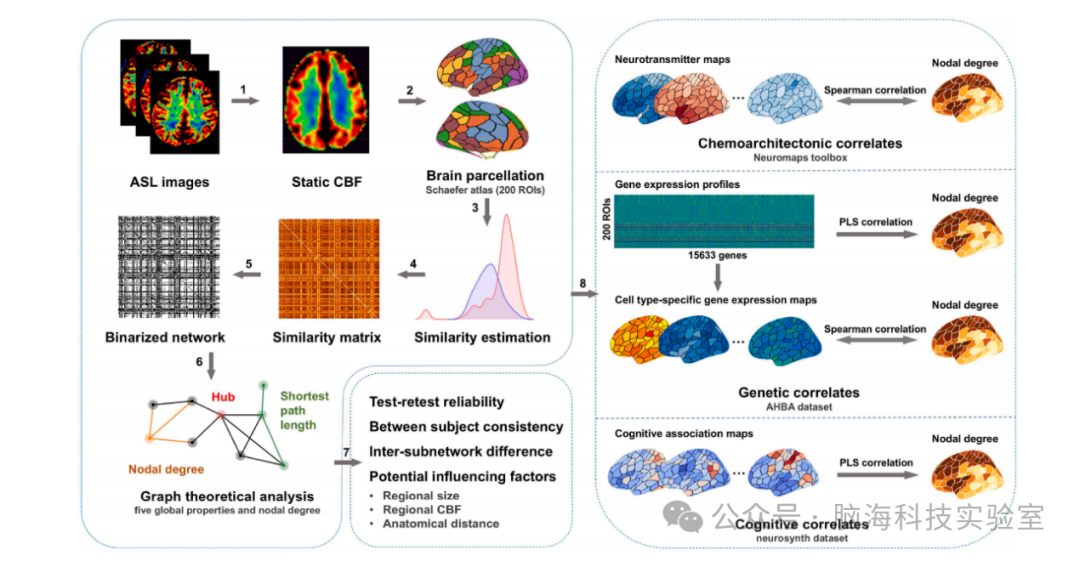
图1 本研究的数据分析流程图。(1)首先对个体的ASL图像进行处理以生成静态CBF图。(2)根据Schaefer图谱将CBF图划分为200个感兴趣区域(ROIs)。 (3)针对每个ROI,基于其中体素的CBF值获得概率分布。(4)对于每对ROI,基于Jensen-Shannon散度估计它们概率分布的相似性。(5)采用基于稀疏性的程序对得到的CBF矩阵进行阈值化,生成一系列二值网络。(6)计算基于图论的网络指标以表征二值网络的拓扑结构。(7)针对区域间CBF相似性和/或CBF网络的拓扑属性,开展分析以评估其重测信度(TRT reliability)、被试间一致性、潜在影响因素以及高阶和低阶亚网络之间的差异。(8)通过将网络的节点度与公开可用的神经递质、基因表达和认知关联图进行相关性分析,探讨CBF网络的化学结构、遗传和认知相关性。ASL:动脉自旋标记;CBF:脑血流;ROIs:感兴趣区域;AHBA:Allen人脑图谱。
2.3 ASL图像处理
使用GE-AW工作站(ADW 4.7,GE Healthcare)的READY view后处理工具,根据以下公式处理ASL图像以生成个体定量静态CBF图:
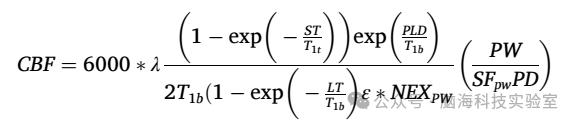
其中,T1b和T1t分别是动脉血(1.6秒)和灰质(1.2秒)的纵向弛豫时间;ST是饱和时间(2秒);λ是分配系数,设置为全脑平均值0.9;ε是反转效率(0.8)和背景抑制效率(0.75)的组合,总效率为0.6;PLD是标记后延迟(1525毫秒);LT是标记持续时间(1500毫秒);PD是参考图像的部分饱和度,使用T1t为1.2秒进行校正;PW表示灌注加权或原始差值图像;SFPW是PW序列的缩放因子;NEXPW是PW图像的激发次数。最终的CBF以mL/100g/min为单位报告。
在进行CBF定量之前,未对ASL图像进行显式头动校正。因此,我们仔细检查了CBF图,并排除了任何显示明显运动伪影的图像,以避免进一步分析。随后,使用ASLtbx工具箱(Hu等,2010;Wang等,2008)对CBF图进行部分体积效应校正,将其与相应的结构图像共配准,归一化到MNI空间,重采样为3毫米各向同性体素,并使用6毫米全宽半高斯核进行空间平滑。最后,通过将每个体素的值除以每个受试者的全脑平均值,对CBF图进行均值缩放,以减少个体间全局CBF变异的影响。值得注意的是,空间平滑和均值缩放均限制在SPM12工具箱中灰质组织概率图(阈值=0.4)导出的灰质掩模内,以排除白质低信噪比的影响。
2.4 CBF网络构建
网络由节点和连接节点的边组成。在本研究中,我们基于每个受试者处理后的定量静态CBF图构建功能脑网络。
节点定义:根据Schaefer图谱(Schaefer等,2018)定义网络节点,该图谱广泛用于皮层分区研究。该图谱将大脑皮层划分为200个感兴趣区(ROIs),这些ROIs进一步被归类为七个亚网络(Thomas Yeo等,2011):视觉网络(VN)、体感运动网络(SMN)、背侧注意网络(DAN)、腹侧注意网络(VAN)、边缘网络(LN)、前顶叶控制网络(FPN)和默认模式网络(DMN)。
边定义:网络边被定义为基于Jensen-Shannon散度(JSD)的区域CBF概率分布的形状相似性(JSDs)。首先,提取每个ROI内的CBF值,并使用MATLAB函数ksdensity估计其概率密度函数。然后将概率密度函数转换为概率分布(PDs),最终通过以下公式定义JSDs:

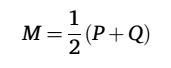
其中,P和Q表示区域概率分布(PDs),n表示概率密度函数估计中使用的采样点数。JSDs的取值范围为[0, 1],值越大表示相似性越高。我们采用与之前研究(Wang等,2016)相同的方法,评估采样点数对区域概率密度函数的影响。具体而言,首先计算每个感兴趣区(ROI)的Fréchet距离,以量化使用不同采样点数(24对25、25对26、26对27、27对28、28对29和29对30)估计的概率密度函数之间的相似性。该分析基于受试者首次扫描的CBF图进行,生成了一个30(受试者)×200(ROI)×6(相邻采样点数对)的Fréchet距离矩阵。随后,该矩阵在受试者间取平均值,得到一个群体水平的Fréchet距离矩阵(200×6)。最后,采用非参数置换检验(重复10000次)比较群体水平Fréchet距离矩阵中任意两列相邻数据。结果显示,随着采样点数的增加,区域概率密度函数的相似性显著且持续增加(p<0.05,通过假发现率(FDR)校正)。这表明更多的采样点能够更稳定地估计基于CBF的区域概率密度函数。为了避免过拟合,并在稳定估计与计算成本之间取得平衡,本研究选择28作为采样点数,与之前的研究一致。
2.5 基于图论的CBF网络拓扑特征分析
我们利用基于图论的网络测量方法对上述构建的CBF网络进行特征分析,所有网络分析均使用GRETNA工具箱完成。
阈值选择。在计算网络指标之前,采用基于稀疏度的阈值化程序将CBF网络二值化,稀疏度定义为实际边数与网络中可能的最大边数之比。基于稀疏度的阈值化程序确保了不同受试者之间边数的一致性。具体而言,每个CBF网络在稀疏度范围[0, 1]内以20个对数间隔点进行阈值化,与我们之前的研究一致。
全局属性。我们在每个稀疏度下计算了五个全局属性:聚类系数(Cp)、最短路径长度(Lp)、局部效率(Eloc)、全局效率(Eglob)和模块化(Q)。为了检验CBF网络是否具有非随机组织特征,我们将所有全局属性进一步标准化,即将其除以100个匹配随机网络的相应平均值。这些随机网络通过拓扑重连算法生成,以保留与CBF网络相同的度分布。一般来说,小世界、高效率和模块化的网络应满足以下条件:标准化Cp > 1且标准化Lp ≈ 1,标准化Eloc > 1且标准化Eglob ≈ 1,标准化Q > 1。考虑到网络属性对稀疏度的依赖性,我们进一步计算了曲线下面积(AUC),为每个全局网络指标提供了一个总结性标量。
节点度。我们在每个稀疏度下计算了每个二值网络的节点度,以表征单个节点在CBF网络中的作用。为了为每个节点提供一个总结性标量,我们还计算了节点度的AUC。随后,将所有受试者的节点度AUC取平均值,并用于拟合度分布模型(幂律、指数和截断幂律模型)。枢纽节点被定义为节点度(AUC)排名在前10%的节点。
2.6 CBF网络的评估
重测信度(TRT reliability)我们使用组内相关系数(Intraclass Correlation Coefficient, ICC)来量化CBF网络的重测信度。具体而言,我们分别计算了每个全局网络指标(AUC)、每个区域的节点度(AUC)以及每对区域间CBF相似性的ICC。对于给定的指标,ICC的计算公式为:
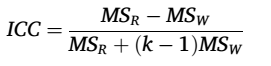
具体而言,MSR表示被试间方差均方,MSW表示被试内方差均方,k表示每个被试的重复测量次数(本研究中为10)。与我们之前的研究类似,我们将重测信度分为以下几类:差(ICC < 0.25)、低(0.25 < ICC < 0.4)、中等(0.4 < ICC < 0.6)、好(0.6 < ICC < 0.75)和优秀(0.75 < ICC < 1)。
被试间一致性(Between-subject consistency)。为了评估CBF网络的被试间一致性,我们计算了每对被试在节点度(跨节点)和CBF相似性矩阵(跨边)上的皮尔逊相关系数,并对所有扫描进行了平均。
亚网络间差异(Inter-subnetwork differences)。为了研究CBF网络是否与皮层的功能拓扑结构相关,我们使用两样本t检验比较了低阶亚网络(视觉网络VN和体感运动网络SMN)与高阶亚网络(背侧注意网络DAN、腹侧注意网络VAN、前顶叶控制网络FPN和默认模式网络DMN)在节点度和亚网络内区域间CBF相似性上的差异。t检验的显著性水平通过非参数置换检验(10000次)估计。同样,我们还比较了低阶和高阶亚网络在节点度的重测信度和亚网络内区域间CBF相似性上的差异。
潜在影响因素(Potential influencing factors)。对于所有被试和扫描平均后的节点度,我们使用斯皮尔曼相关系数(Spearman correlation)分析了它们与区域大小和平均CBF水平的关系。这些相关性的显著性水平通过旋转测试(spin tests)估计。旋转测试是一种基于皮层表面球面投影角度置换的空间置换方法,它保留了数据的空间协方差结构,从而校正了空间自相关的潜在混杂效应。具体而言,我们首先生成了10000次平均节点度图(即源图)的随机空间旋转(即旋转),并将每次旋转与目标图(即区域大小图或平均CBF图)进行相关性分析。然后,计算P值,即旋转后相关系数的绝对值大于或等于源图与目标图实际观察到的相关系数绝对值的比例。此外,我们还使用斯皮尔曼相关系数分析了所有被试和扫描平均后的区域间CBF相似性与区域间解剖距离(欧几里得距离)的关系。
2.7 CBF网络的化学结构相关性研究
为探究CBF网络的化学结构相关性,我们首先从链接(Hansen等,2022)获取了38名健康受试者的神经递质强度图。这些强度图涵盖了以下神经递质受体和转运体:5-羟色胺受体1A(5-HT1A)、5-HT1B、5-HT2A、5-HT4、5-HT6、5-羟色胺转运体(5-HTT)、α4β2烟碱型乙酰胆碱受体(α4β2)、大麻素受体1(CB1)、多巴胺受体1(D1)、D2、多巴胺转运体(DAT)、γ-氨基丁酸A型受体(GABAA)、组胺H3受体(H3)、毒蕈碱型乙酰胆碱受体1(M1)、代谢型谷氨酸受体5(mGluR5)、μ型阿片受体(MOR)、去甲肾上腺素转运体(NET)、N-甲基-D-天冬氨酸受体(NMDA)以及囊泡乙酰胆碱转运体(VAChT)。由于使用raclopride示踪剂的D2强度图在皮层中的结合不可靠,因此将其排除在后续分析之外。关于这些神经递质强度图的详细信息可参考链接(Markello等,2022)。
我们进一步分析了所有受试者和扫描中节点度的平均值与每种神经递质的区域平均强度之间的关系,采用Spearman相关性分析(针对节点)。通过基于旋转的置换检验(10,000次)评估这些相关性的显著性水平,并采用假发现率(FDR)程序对所有相关性进行多重比较校正,校正水平为q < 0.05。
2.8 CBF网络的遗传相关性研究
为探究CBF网络的遗传相关性,我们研究了所有受试者和扫描中节点度的平均值与艾伦人脑图谱(AHBA)数据集中的转录组特征之间的关系。对于在该关系中贡献显著的基因,我们进一步进行了基因本体(GO)富集分析,以识别其相关的生物学过程。
AHBA数据集。AHBA数据集是一个公开的在线资源,包含来自6名健康成年供体(年龄24-57岁;5名男性和1名女性)的全脑转录组信息,这些供体均无已知的神经病理学或神经精神疾病史(Hawrylycz等,2012)。具体而言,该数据集记录了来自3702个空间上不同的脑组织样本的20,737个基因的转录活性,这些样本几乎覆盖了整个大脑。其中,4名供体的样本取自左侧大脑半球,而2名供体的样本取自双侧大脑半球。更多细节请参考艾伦人脑图谱官方网站(该链接目前无法访问,可能需要检查拼写或稍后重试)。基因数据的预处理采用了标准化流程(Arnatkevičiūtė等,2019),具体方法见补充方法部分。
CBF网络与转录组特征之间的关系。为研究CBF网络与转录组特征之间的关系,我们采用偏最小二乘回归(PLS)方法,以所有基因的区域表达水平预测所有受试者和扫描中节点度的平均值。PLS回归的第一个成分(PLS1)是所有基因区域表达水平的线性组合,与节点度的相关性最强。通过基于旋转的置换检验(10,000次)重新运行PLS回归,模拟节点度,以校正空间自相关,并评估相关性的显著性水平。如果相关性显著,则对形成PLS1的所有基因权重进行Z转换,绝对Z值大于1.64的基因被认为对相关性贡献显著。对于这些对PLS1贡献显著的基因,我们进一步进行了基因本体(GO)富集分析,以寻找其相关的GO术语(具体方法见补充部分)。值得注意的是,GO富集分析分别针对对PLS1贡献最强的正相关基因(PLS1+)和负相关基因(PLS1-)进行。
此外,为探讨CBF网络的节点度是否与特定细胞结构的基因表达相关,我们计算了节点度与以下细胞类别的富集基因的平均表达水平之间的Spearman相关性,这些细胞类别包括:兴奋性神经元、抑制性神经元、少突胶质细胞前体细胞、星形胶质细胞、内皮细胞、小胶质细胞和少突胶质细胞(Arnatkeviciute等,2021)。所有相关性的显著性水平通过基于旋转的置换检验(10,000次)进行估计,并采用假发现率(FDR)程序对所有细胞类别进行多重比较校正(校正水平为q < 0.05)。
2.9 CBF网络的认知相关性研究
为研究CBF网络的认知相关性,我们采用偏最小二乘回归(PLS)分析,将所有受试者和扫描中节点度的平均值与多个基于元分析的激活图进行相关性分析。这些激活图来源于NeuroSynth数据库(Yarkoni等,2011)https://github.com/neurosynth/neurosynth,该数据库反映了大脑体素与神经认知过程术语之间的关联,即在特定体素观察到神经激活时,某术语在脑成像实验中被报告的概率。具体而言,我们基于认知图谱(Cognitive Atlas)(Poldrack等,2011)选择了123张主要与认知功能相关的激活图,其完整列表可在Hansen等(2022)的研究中找到。
这些激活图通过Schaefer脑图谱进行分割,并进行Z转换以获得区域激活评分,用于预测CBF网络的节点度。与前面类似,我们仅考察了PLS回归的第一个成分(PLS1),其显著性水平通过基于旋转的置换检验(10,000次)进行估计(Alexander-Bloch等,2018;Vaša等,2018)。如果相关性显著,我们进一步计算了PLS1评分与每张激活图的区域激活评分之间的皮尔逊相关性。这些相关性的显著性水平同样通过基于旋转的置换检验(10,000次)进行估计,并采用Bonferroni校正方法对所有相关性进行多重比较校正(校正水平为p < 0.05/123)。
3 结果
3.1 CBF网络的连接性和拓扑组织
连接模式。图2a展示了所有受试者和扫描中平均化的CBF相似性矩阵。大脑区域之间的相似性较高(0.708 ± 0.096)。
全局组织。每个CBF网络在大多数阈值范围内表现出小世界特性、高效率和模块化结构,这通过归一化的聚类系数(Cp)>1、归一化的路径长度(Lp)≈1、归一化的局部效率(Eloc)>1、归一化的全局效率(Eglob)≈1以及归一化的模块化系数(Q)>1来证明(图2b)。
中心节点(Hub)。CBF网络的度分布最适合指数截断幂律模型(R² = 0.985)(图2c)。共识别出20个中心节点区域,主要分布在默认模式网络(DMN,5个区域)、背侧注意网络(DAN,5个区域)、感觉运动网络(SMN,4个区域)和视觉注意网络(VAN,4个区域)(图2d)。
3.2 CBF网络的可靠性与一致性
CBF网络的全局网络指标表现出较低的重测信度(Test-Retest Reliability, TRT):聚类系数(Cp:ICC = 0.347)、路径长度(Lp:ICC = 0.200)、局部效率(Eloc:ICC = 0.345)、全局效率(Eglob:ICC = 0.277)、模块化系数(Q:ICC = 0.259)以及归一化的相应指标(归一化Cp:ICC = 0.222;归一化Lp:ICC = 0.249;归一化Eloc:ICC = 0.221;归一化Eglob:ICC = 0.203;归一化Q:ICC = 0.314)。
对于节点度,尽管总体重测信度较低(ICC = 0.210 ± 0.104),但特定一组区域(9个,占比4.5%)表现出中等的重测信度(图3a)。这些区域主要位于视觉注意网络(VAN,4个区域,占比44.4%)和默认模式网络(DMN,3个区域,占比33.3%),例如右颞-枕-顶交界区和左前额叶皮质。同样,尽管区域间CBF相似性的总体重测信度较低(ICC = 0.206 ± 0.102),但特定一组连接(888条,占比4.46%)表现出中等的重测信度(图3b)。
对于被试间一致性,节点度(r = 0.746 ± 0.103)(图3c)和区域间CBF相似性(r = 0.654 ± 0.112)(图3d)均表现出较高的被试间一致性。
3.3 CBF网络的亚网络间差异
高阶亚网络的内网络CBF相似性显著高于低阶亚网络(t = 7.740, p < 0.001)(图4a)。此外,与低阶亚网络相比,高阶亚网络的内网络CBF相似性表现出显著较低的重测信度(t = -9.210, p < 0.001)(图4b)。在低阶和高阶亚网络之间,节点度(t = 0.860, p = 0.383)以及节点度的重测信度(t = 1.259, p = 0.206)均未观察到显著差异。
3.4 CBF网络的影响因素
CBF网络的节点度与区域大小显著负相关(rho = -0.339, p < 0.001)(图5a),但与区域平均CBF水平显著正相关(rho = 0.601, p < 0.001)(图5b)。对于区域间CBF相似性,其与区域间解剖距离显著负相关(rho = -0.239, p < 0.001)(图5c)。
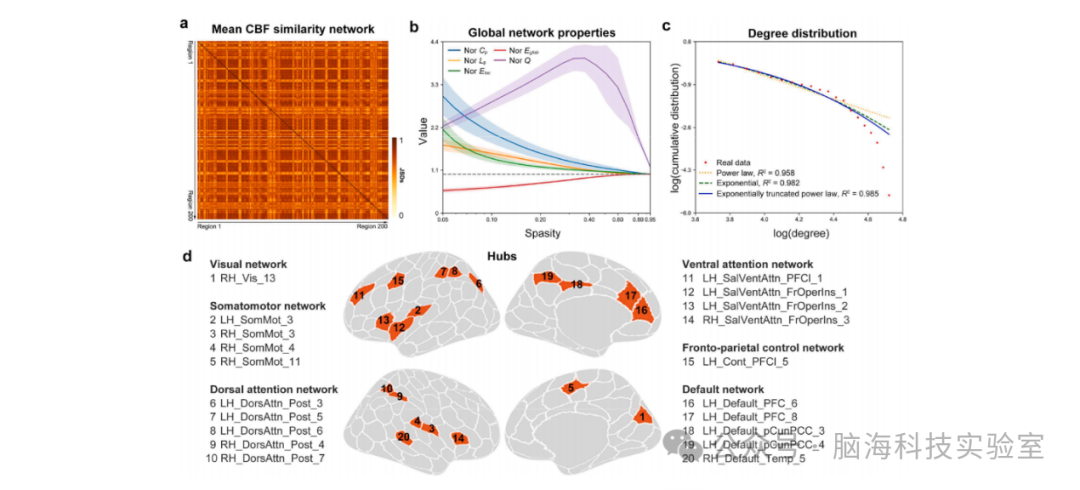
图2 CBF网络的拓扑组织。(a) 群体水平的平均CBF相似性矩阵。(b) CBF网络表现出小世界特性、高并行效率和模块化结构。(c) CBF网络的度分布最适合指数截断幂律模型。(d) 共识别出20个中心节点区域。CBF:脑血流;JSDs:基于Jensen-Shannon散度的相似性;Nor Cp:归一化聚类系数;Nor Lp:归一化最短路径长度;Nor Eloc:归一化局部效率;Nor Eglob:归一化全局效率;Nor Q:归一化模块化系数;LH:左侧大脑半球;RH:右侧大脑半球;FrOper:额叶盖部;Ins:岛叶;PFCl:外侧前额叶皮质;pCunPCC:楔前叶/后扣带回皮质;Temp:颞叶
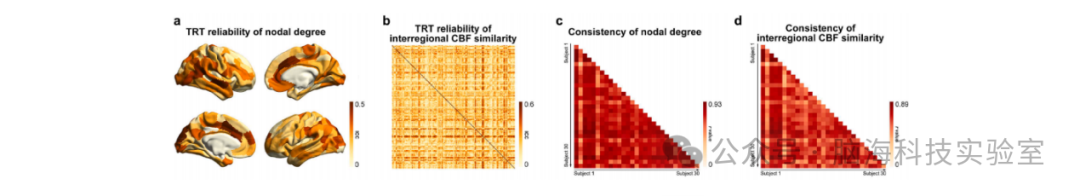
图3 CBF网络的重测信度和被试间一致性。CBF网络在节点度(a和c)以及区域间CBF相似性(b和d)方面表现出较低到中等的重测信度和较高的被试间一致性。TRT:重测信度;CBF:脑血流(Cerebral Blood Flow);ICC:组内相关系数。
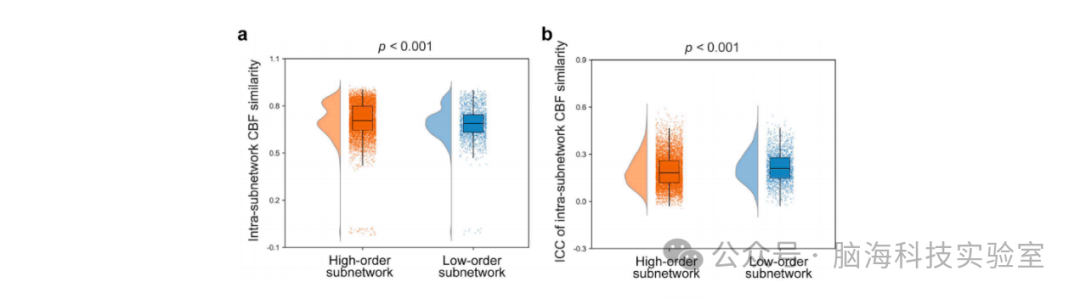
图4 低阶子网络与高阶子网络在(a)区域间CBF相似性和(b)其重测信度(TRT)方面的差异。与低阶子网络相比,高阶子网络表现出显著更高的区域间CBF相似性,但这些相似性的重测信度较低。CBF:脑血(Cerebral Blood Flow)。
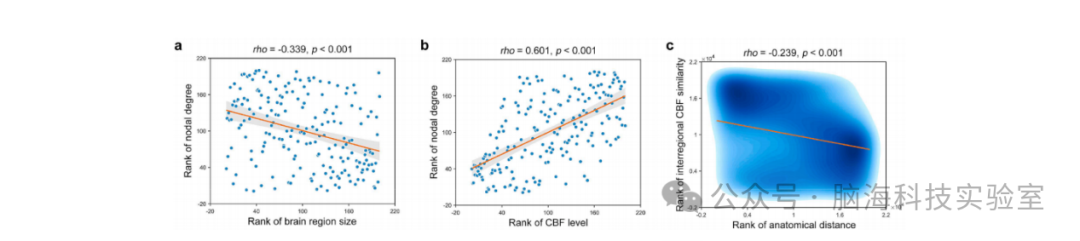
图5 影响CBF网络的因素。(a) CBF网络的节点度与区域大小呈显著负相关。(b) CBF网络的节点度与区域脑血流(CBF)水平呈显著正相关。(c) 区域间CBF相似性与区域间解剖距离呈显著负相关。所有相关性均基于非参数斯皮尔曼相关性分析,因此显示了相应变量的秩次。CBF:脑血流(Cerebral Blood Flow)。
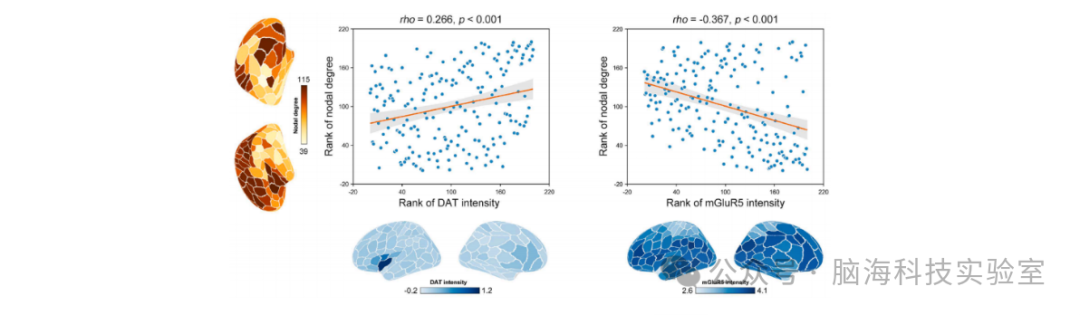
图6 CBF网络的化学结构相关性。CBF网络的节点度与多巴胺转运体(DAT)的区域强度呈显著正相关,与代谢型谷氨酸受体5(mGluR5)的区域强度呈显著负相关。所有相关性均基于非参数斯皮尔曼相关性分析,因此显示了相应变量的秩次。DAT:多巴胺转运体;mGluR5:代谢型谷氨酸受体5。
3.5 CBF网络的化学结构相关性
CBF网络的节点度与多巴胺转运体(DAT)的区域强度呈显著正相关(rho = 0.266, p < 0.001),使用[11C]P943示踪剂;同时与代谢型谷氨酸受体5(mGluR5)的区域强度呈显著负相关(rho = -0.367, p = 0.002),使用[11C]ABP688示踪剂(图6)。其他神经递质图的强度未观察到显著相关性(p > 0.05,经假发现率校正)。
3.6 CBF网络的遗传相关性
CBF网络的节点度与基于区域基因表达谱的PLS1评分呈显著正相关(r = 0.530, p = 0.043)(图7a)。共识别出427个正相关基因和383个负相关基因(图7b)。基因本体(GO)富集分析显示,PLS1+基因富集于17个生物学过程,主要涉及胆固醇相关过程(如“细胞内胆固醇运输”和“胆固醇稳态”)以及内皮细胞相关过程(如“内皮细胞凋亡的正向调控”和“内皮细胞分化的调控”)(附表S2)。PLS1-基因未观察到神经生物学过程。细胞类型特异性分析显示,CBF网络的节点度与内皮细胞富集基因的平均表达水平呈显著正相关(rho = 0.332, p < 0.001)(图7c),其他细胞类型未观察到显著相关性(p > 0.05,经假发现率校正)。
3.7 CBF网络的认知相关性
我们观察到CBF网络的节点度与基于123张元分析激活图的PLS1评分呈显著正相关(r = 0.573, p = 0.003)(图8a)。PLS1评分与24张激活图呈显著正相关,与28张激活图呈显著负相关(p < 0.05,经Bonferroni校正)。相关性最高的正相关激活图主要涉及执行功能(如“目标”、“监控”、“干扰”、“反应选择”和“计划”)和“工作记忆”,而相关性最高的负相关激活图主要与语言领域相关,如“交流”、“含义”、“倾听”、“语言理解”、“言语感知”和“语言”(图8b)。
4讨论
在本研究中,我们提出了一种通过估计区域静态脑血流(CBF)概率分布的形状相似性来构建功能脑网络的方法。我们系统地评估了构建的CBF网络,分析了其拓扑组织、重测信度(TRT)、被试间一致性、与皮层功能拓扑的对齐性、影响因素以及与化学结构、基因表达和认知功能的关联。我们发现,CBF网络表现出非平凡的组织特性、较低到中等的重测信度以及较高的被试间一致性。我们进一步证明了CBF网络与区域大小和CBF水平相关,依赖于解剖距离,并受到皮层功能拓扑的调节。此外,我们还发现CBF网络与多巴胺转运体(DAT)和代谢型谷氨酸受体5(mGluR5)强度图共变,与富集于胆固醇和内皮细胞相关过程的基因相关,并编码语言、记忆和执行功能。总体而言,这些发现表明,区域间静态CBF概率分布的形状相似性为研究功能脑网络提供了一种新颖、可靠且具有神经生物学意义的方法,是对当前方法的重要补充。
4.1 CBF网络的非平凡组织特性
我们发现,CBF网络表现出小世界属性,这种特性被认为可以支持人类大脑中信息的高效分离和整合,同时最小化连接成本[Bassett和Bullmore, 2006;Liao等, 2017]。小世界属性在其他类型的脑网络中也经常被观察到,例如基于BOLD的功能网络[Wang等, 2009a]、结构网络[Hagmann等, 2007]和形态学网络[Li等, 2021a;Wang等, 2016]。与以往研究一致,我们的发现进一步支持了小世界属性是人类脑网络的普遍组织原则的观点。
除了小世界属性外,我们还发现CBF网络表现出高度的模块化,表明CBF网络可以被划分为几个不同的、相互作用的模块。模块化通过将某些功能定位到特定模块,同时保持模块间通信,从而在网络中实现高效的信息处理[Bullmore和Sporns, 2009;Meunier等, 2010]。因此,模块化可能提高了人类脑网络的稳健性、适应性和进化能力,并支持适应性行为和认知的出现[Bullmore和Sporns, 2009;Meunier等, 2010]。
最后,我们在CBF网络中识别出几个中心节点区域,并发现CBF网络的节点度与区域CBF水平呈显著正相关。这些发现表明,CBF水平较高的区域倾向于成为CBF网络的中心节点,这与基于BOLD的功能脑网络的结果一致[Liang等, 2013b]。以往研究表明,CBF水平较高的区域通常与增强的神经活动、密集的毛细血管网络以及对氧和葡萄糖的高能量需求相关[Karbowski, 2011;Watts等, 2018]。这些特征可能使它们参与更多的神经处理,从而成为功能脑网络的中心节点。
4.2 CBF网络的重测信度
在本研究中,我们评估了CBF网络的重测信度和被试间一致性。我们观察到全局网络指标、节点度和区域间CBF相似性的重测信度较低到中等。以往研究表明,CBF受参与者身体和情绪状态的影响[Honda等, 2018;Hoshi和Chen, 2002;Ozawa等, 2019]。此外,CBF还表现出明显的昼夜节律变化[Hodkinson等, 2014;Shannon等, 2013],以及在女性月经周期中的变化[Otomo等, 2020]。然而,这些因素在数据收集阶段未被考虑,推测它们在一定程度上共同导致了CBF网络的低到中等的重测信度。在未来的研究中,研究这些因素对通过我们提出的方法构建的CBF网络的影响将是非常有意义的。
除了这些因素外,不同的图像预处理策略(例如,运动校正和空间滤波)也被报道会影响CBF的重测信度[Fazlollahi等, 2015],这也可能是导致CBF网络低到中等重测信度的原因之一。解析不同图像预处理策略对CBF网络重测信度的影响,将为确定获得可靠CBF网络的优化分析策略提供指导。
尽管CBF网络的重测信度较低到中等,但我们观察到CBF网络在被试间具有较高的被试间一致性。这种高一致性表明存在一种稳定的、内在的CBF网络模式,这种模式在被试间是共享的。因此,我们对个体CBF网络进行平均,以得出一个共识估计值,该估计值被用于探索CBF网络的神经生物学特征。
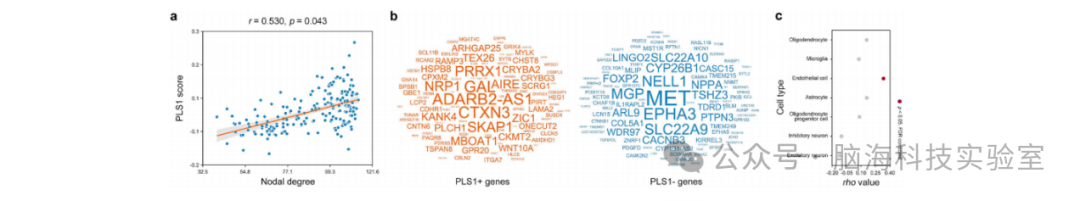
图7 CBF网络的遗传相关性。(a) CBF网络的节点度与基于15,633个基因转录活性的PLS1评分呈显著正相关。(b) 共识别出427个基因对PLS1评分呈正贡献,383个基因呈负贡献。(c) CBF网络的节点度与内皮细胞富集基因的平均转录活性呈显著正相关。PLS:偏最小二乘回归;FDR:假发现率。
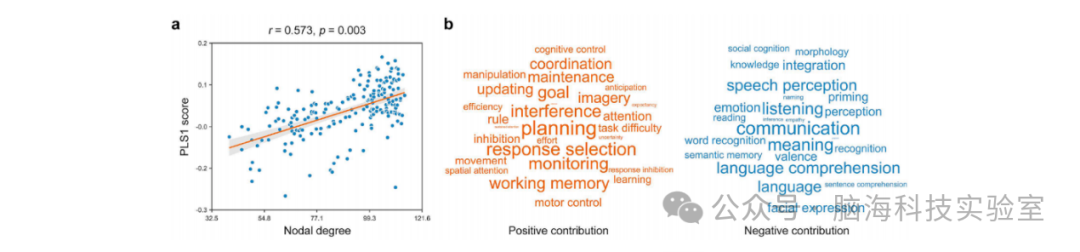
图8 CBF网络的认知相关性。(a) CBF网络的节点度与基于123张激活图的PLS1评分呈显著正相关。(b) 共识别出24个认知过程对PLS1评分呈正贡献,28个认知过程呈负贡献。PLS:偏最小二乘回归。
4.3 CBF网络的神经生物学特征
我们通过将CBF网络与神经递质强度和转录组特征联系起来,探讨了其神经生物学特征。我们发现CBF网络的节点度与多巴胺转运体(DAT)强度呈显著正相关,而与代谢型谷氨酸受体5(mGluR5)强度呈负相关。以往研究显示,不同精神类药物诱导的CBF变化与神经递质系统相关,包括DAT[Dukart等, 2018]。关于mGluR5,其激活被发现可触发星形胶质细胞中的Ca²⁺瞬变[Wang等, 2006],这一过程通过调节小动脉平滑肌张力最终影响CBF[Zonta等, 2003]。因此,CBF网络与DAT和mGluR5强度的关联可能由这些神经递质对CBF动态的调节作用所驱动。
除了化学结构相关性外,我们还发现CBF网络与转录组特征显著相关。此外,我们发现对这种相关性贡献显著的基因主要富集于胆固醇相关过程,包括细胞内胆固醇运输和胆固醇稳态。生理学上,胆固醇在维持最佳能量代谢水平中发挥关键作用[Czuba等, 2017]。考虑到CBF与葡萄糖和氧代谢密切相关[Hoge等, 1999;Paulson等, 2010;Vaishnavi等, 2010b],我们推测胆固醇可能通过调节能量需求进而影响大脑中的CBF分布,从而进一步影响CBF网络的拓扑组织。鉴于我们观察到的区域CBF与CBF网络节点度之间的显著正相关,这一推测似乎合理。除了胆固醇相关过程外,PLS1+基因还富集于多个内皮细胞相关过程,例如内皮细胞凋亡的正向调控和内皮细胞分化的调控。这些发现与我们的细胞类型特异性分析结果一致,后者揭示了CBF网络的节点度与内皮细胞富集基因的平均表达水平呈显著正相关。以往多项研究报道了内皮细胞在CBF调节中的作用,包括血管张力调节、炎症反应、血栓形成、黏附和血管通透性[Ashby和Mack, 2021;Cohen, 1995;Godo和Shimokawa, 2017]。在此,我们的结果表明内皮细胞可能也在CBF网络的拓扑组织中发挥关键作用。综上所述,这些发现为理解通过我们提出的方法构建的CBF网络的神经生物学基础提供了重要见解。
4.4 CBF网络的认知和行为相关性
在本研究中,我们通过将CBF网络与一系列元分析激活图联系起来,证明了其认知和行为相关性。我们发现CBF网络主要与执行功能、工作记忆和语言处理相关。这些发现与认知源于分布式大脑系统在大规模网络中的功能交互的观点一致[Bressler和Menon, 2010]。值得注意的是,以往研究发现较高的区域CBF与更好的执行功能和记忆表现相关[Leeuwis等, 2018]。因此,认知表现似乎不仅与区域CBF相关,还与CBF网络的拓扑组织相关。在未来的研究中,有必要探讨CBF网络是否能够解释个体间的差异,并预测个体在行为和认知中的得分。特别是考虑到CBF网络的高度模块化,识别CBF网络的模块组成,并确定不同模块如何与多样的行为和认知功能具体相关,将是一个有趣的方向。
4.5 限制与未来方向
本研究存在一些限制。首先,动脉自旋标记(ASL)成像技术存在固有问题,例如血管伪影。尽管在ASL数据采集过程中使用了血管压制技术,我们仍不确定结果是否仍受到血管伪影的干扰。因此,未来研究需要通过使用具有多种标记持续时间和/或标记后延迟的ASL数据来检验我们方法的稳健性,这些数据能够考虑可变动脉传输时间对脑灌注准确评估的影响[Alsop等, 2015;Woods等, 2024]。其次,我们通过将CBF网络与公开可用的数据集联系起来探讨其生物学意义,这可能导致对CBF网络的化学结构和遗传相关性的低估。因此,需要进一步工作,通过从同一队列的参与者中收集所有数据来验证我们的结果。第三,目前有多种使用CBF构建功能脑网络的方法[Liang等, 2014b;Liu等, 2016]。因此,将本研究提出的方法与这些现有方法进行比较,以更好地理解其优缺点,将是非常重要的。第四,以往研究表明,不同的网络分析策略对人类脑网络的拓扑组织有深远影响[Brodoehl等, 2020;Liang等, 2012;Wang等, 2009b;Zalesky等, 2010]。因此,未来研究需要探索在使用不同的脑区划分、阈值方法和网络类型时,我们的结果是否具有可重复性。第五,我们使用静态CBF构建功能脑网络,因为区域CBF与神经活动高度相关,这是由于神经-血管耦合。然而,以往研究发现,CBF反应的动态范围小于BOLD反应在视觉刺激亮度对比中的动态范围[Liang等, 2013a]。因此,与基于BOLD的功能连接方法相比,我们提出的方法可能在揭示外部刺激引发的功能脑网络重组方面不够敏感。因此,未来研究需要检验这两种方法得出的功能脑网络之间的相似性和差异性。最后,静态CBF的变化已在多种神经精神疾病中被发现[Falcon等, 2024;Li等, 2021a;Percie Du Sert等, 2023]。因此,未来一个有趣的研究方向是探讨通过我们提出的方法构建的CBF网络是否也在这些疾病中受到破坏。
数据和代码可用性声明
本研究中支持研究结果的所有数据均来自公开数据集(CBF数据集:https://fcon_1000.projects.nitrc.org/indi/CoRR/html/hnu_1.html;神经递质受体和转运体数据集:https://github.com/netneurolab/neuromaps;元分析激活数据集:https://neurosynth.org/)。本研究中所有分析的代码均来自公开工具箱,包括GRETNA(https://www.nitrc.org/projects/gretna;拓扑分析)、abagen(版本0.1.3;https://github.com/rmarkello/abagen;基因数据预处理)以及GeneCategoryEnrichmentAnalysis(https://github.com/benfulcher/GeneCategoryEnrichmentAnalysis;基因本体富集分析)。
补充材料
与本文相关的补充材料可在在线版本中找到,DOI为:10.1016/j.neuroimage.2025.121069。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









