






目录
1研究背景
传统静息态fMRI研究表明,大脑活动具有时空约束性(如静息态网络,RSNs),但动态功能网络(dFC)的生成机制尚不明确。现有方法(如HMM、CAP)仅能捕捉有限的时间动态特征,缺乏统一框架解释复杂时空模式。核心问题在于如何从非线性、非平稳的fMRI信号中提取基础时间结构。为此,研究人员提出局部空间对比学习(Local Space-Contrastive Learning, LSCL),该研究为一种新型非线性空间独立成分分析(sICA)方法,通过无监督深度学习提取重复出现的局部时间基元(temporal primitives),以统一解释静息态与任务态fMRI的动态机制。
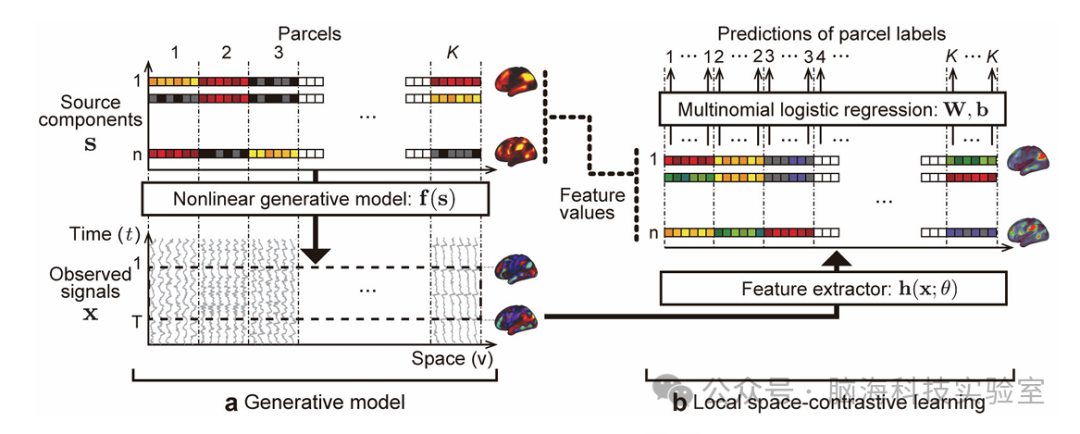
图1.局部空间对比学习(LSCL)的概念
2研究方法
2.1 LSCL框架设计
核心思想:假设fMRI信号由空间非平稳、局部非线性时间基元生成,通过训练深度时间卷积神经网络(CNN)提取这些时间基元。
关键步骤:①空间分割:将皮层划分为1,833个功能分区(parcels)。②局部时间窗口的构建:将每个分区的时间序列划分为多个短时间窗口。③特征提取:使用CNN处理短时间窗口(46个时间点,约32秒)的fMRI时间序列,通过多分类逻辑回归(MLR)预测每个时间片段所属的功能分区,迫使网络学习空间特异性的非线性时间模式。③线性ICA后处理:将CNN提取的特征值输入线性ICA,以消除特征间的线性不确定性。
2.2数据与预处理
数据来源:HCP项目的1,003名受试者静息态和任务态fMRI数据。
预处理:ICA-FIX去噪,仅保留皮层表面顶点数据,时间序列标准化。
2.3验证与分析
时间基元表征:通过时间对齐和频谱分析提取代表性模式。
空间共现模式:t-SNE可视化时间基元的空间共现网络。
任务关联性:分析时间基元在任务态中的时序和空间分布。
行为关联分析:计算时间基元活动与流体智力、工作记忆等行为特征的相关性。
3研究结果
3.1识别五类时间基元
C1:0.13 Hz振荡模式(可能与记忆巩固相关);
C2:瞬态血流动力学响应(HRF)和呼吸响应函数(RRF),驱动RSN动态共现,与神经活动相关;
C3:负BOLD响应(NBR),与额顶控制网络(FPN)相关;
C4:任务持续性BOLD活动(如DAN、FPN),与任务相关BOLD响应的持续期有关;
C5:高频噪声(如呼吸/心跳伪影),可能与生理噪声有关。
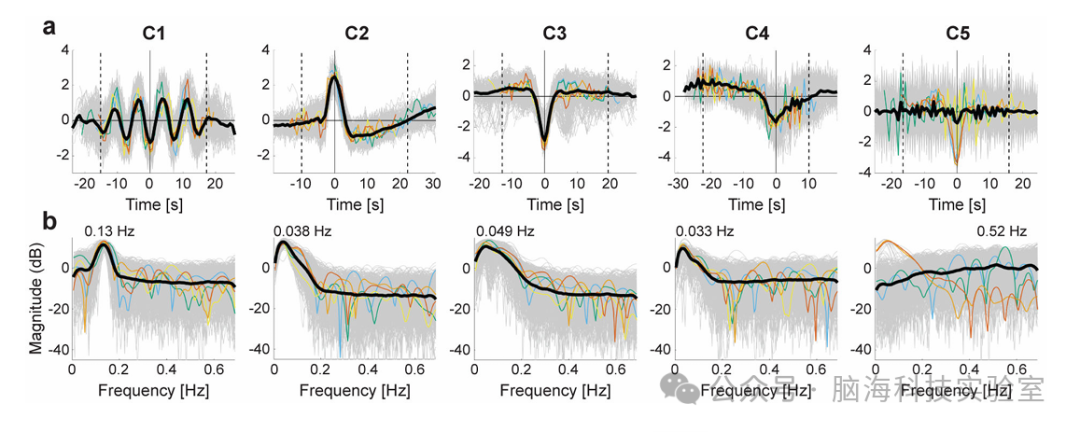
图2.(a)时间基元的代表性时间模式(最重要的时间特征)的可视化,这些基元是由非线性特征提取器(CNN)学习的特定于组件的时空局部非线性时间结构。它们通过取每个组件维度上(未混合的)特征值非常大的时间片段的平均时间来表示。灰色细线是在整个数据集中产生前0.0001%最高组件活动的单个输入时间序列。彩色细线表示激活度最高的样本(第1至5个:红色、橙色、黄色、绿色和蓝色)。考虑到CNN 的移位不变性(进一步评估见图 4a),所有样品都进行了时间偏移,以最大限度地提高它们与参考信号的互相关,即具有最高活性的信号(红色样品)。黑色粗线显示时间偏移后的样本平均值。两条垂直虚线表示颞窗的边缘,其宽度与特征提取器的感受野宽度相同;选择窗口内的(绝对)峰点作为参考点。可以看到,窗口内的平均时间模式在组件之间显示出明显的差异,并在此后被用作时间基元的代表性时间模式。(b)时间基元的代表性频谱。对 a 中的每个 shift 调整输入的频谱进行了估计。与a一样,灰色细线是单个图,彩色细线表示活化度最高的样本,黑色粗线表示它们的平均值。平均频谱的峰值频率显示在线上。
3.2时间基元的时空特性
图3a以直方图形式展示了时间基元的出现频率(特征值分布)情况。特征值反映了时间基元在数据中出现的频率和强度。与通过自回归随机化(ARR)生成的代理数据的特征值分布相比,实际数据中特征值显著高于随机水平,具体超出比例分别为0.28%、1.4%、1.3%、0.10%和0.43%,且均超越了未校正的阈值α=0.001。这一结果揭示了这些时间片段中存在与时间基元相关的时间结构。不同时间基元(C1至C5)的特征值分布显示出显著差异,表明它们在数据中出现的频率和模式不同。尤其C2和C3的实现比例相较于其他成分异常突出,表明它们在fMRI数据中更为常见。代理数据对时间基元的敏感性差异进一步凸显了时间基元的某些独特属性。例如,与真实数据相比,随机块状替代数据更强烈地激活了C5,这似乎表明C5专注于捕获数据中的高频噪声,而即便是C1,尽管也代表了相对较高的频率时间模式(如图2所示),也呈现出不同的激活特征。
图3b展示了时间基元在大脑不同区域的显著性比率,清晰展现了不同时间基元的空间非平稳性,这与LSCL的预设假设相契合。尽管存在大量重叠区域,但各时间基元的分布特征仍有所不同。特别是,C1、C2和C3在视觉皮层、下顶叶(IPL)以及中间颞回(MTG)的部分区域呈现出相似的分布模式。然而,它们之间也存在一些细微差异:C1在视觉皮层和颞叶区域的出现频率较高。C2在感觉运动区和视觉皮层的出现频率较高,而C3在顶叶和枕叶区域的出现频率较高。C4的分布范围相对更广,其模式似乎融合了任务正网络和默认模式网络(DMN)的特点。C5则在噪声敏感区域(如下颞叶)表现出较高的出现频率。需注意的是,这些空间分布并不直接反映时间基元的空间共存(即网络)情况(见图6),因为这些分布图是基于每个包裹的实现数量计算得出的,而未考虑其时间上的实现数量(具体过程请见图5说明)。在LSCL框架下,空间共存模式实际上可能呈现出不同的特征。
图3c展示了每个时间基元在不同受试者中的显著性比率的变异性,结果表明时间基元的显著性比率在不同受试者之间存在一定的变异性,但总体上表现出一致的模式。C5在某些受试者中可能几乎不出现。
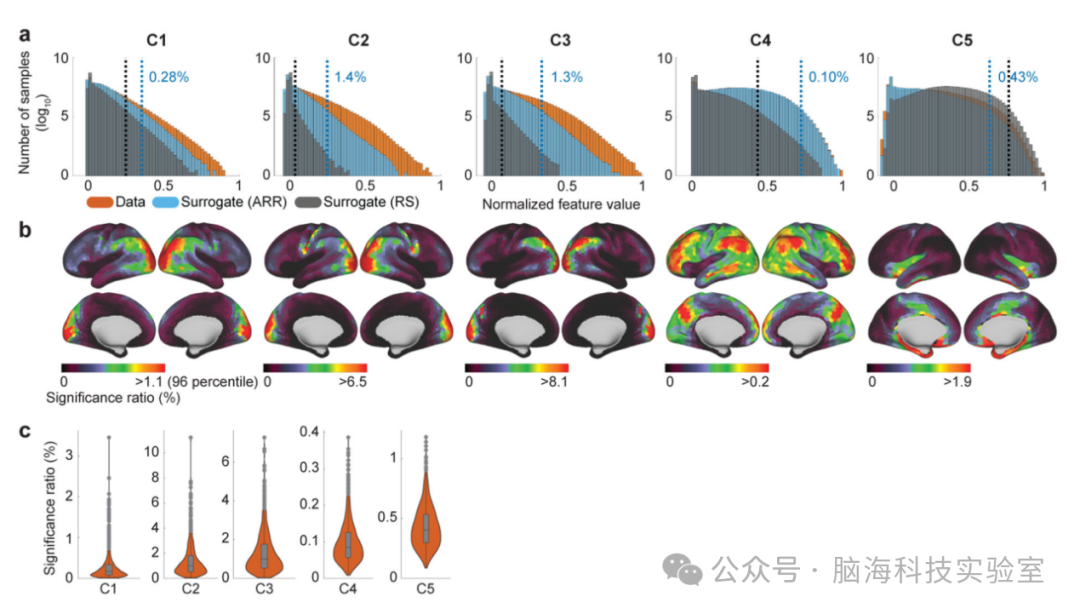
图3.从整个数据集(橙色)获得的实际数据中特征值的直方图(对数刻度),及其与来自代理数据的特征值的比较(蓝色:自回归随机化 (ARR);灰色:随机随机化 (RS))。特征值的尺度被归一化为具有每个组件的最大值 1。蓝色垂直线表示从 ARR 代理数据估计的特征值的几率水平(α = 0.001;未校正)。特征值的 0.28%、1.4%、1.3%、0.10% 和 0.43% 超过随机水平。为了进行比较,黑色垂直线显示根据 RS 代理数据估计的阈值(α = 0.001;未校正)。不同时间基元对代理数据的敏感性的差异表明了时间基元的一些特征;例如,RS 代理数据中 C5 的较高特征值似乎表明它捕获了数据中的高频(可能是生理学)伪影,而这些伪影往往与 RS 代理数据一样缺乏时间结构。(b) 特征值显著性比的位置可视化显示了实现的空间不稳定性和组件特异性。与 a 的几率级别相同。重要的是,正如文中所解释的那样,这些空间分布并不表示共激活网络。(c)不同受试者中的显著性比率的变异性。箱线图中的数据点表示受试者中实现的比率。该框绘制在 25 和 75 个百分位数之间,其中一条线表示中位数。须线表示四分位距的 1.5 倍
3.3行为关联
在本研究的框架中,最基础的非线性特性体现在对时间变化的不变性上。这主要归因于输入样本是从数据矩阵中随机裁剪得到的,无需考虑大脑活动的实际时序。同时, rfMRI数据并未包含任何任务设计。此外,由于实验者采用的CNN具有卷积性质,因此天生具备学习移位不变特征的能力。如图4a所示,时间移位在数据中普遍存在,且各成分从线性回归分析的小斜率中展现出一定的移位不变性。
为了评估时间偏移之外的非线性调制程度,实验者在补偿时间偏移后,分析了特征值与代表模式相似性之间的关系(图4b)。结果显示,特征提取器对类似代表模式的时间模式具有较高的敏感性(C5除外)。当输入较少时,特征提取器会减弱响应,导致特征值的稀疏性(C5除外)。此外,这种关系呈现出非线性特征,与线性模型相比,直方图具有更广泛的水平分布,表明非线性特征提取器具有广泛的调制范围,这得益于其非线性特性。由于C5代表fMRI噪声,实验者在后续分析中将其省略。
为了直观展示实现的非线性调制,实验者将时间基元的实现基于相似性嵌入到二维空间中,并绘制了分布图(图5a)。散点图在特征值与其嵌入位置之间未显示出明显关联,再次证明了特征提取器对特定于成分的模式具有很强的不变性。与线性特征提取器相比,非线性特征提取器不仅关注最大激活的输入模式,还能对偏离该模式的输入进行不同程度的激活。此外,分布图未显示出清晰的聚类结构,表明可变性是非结构性的,不易在组件中进一步分类。这有助于将组件正确划分为结构特定的组件,避免受到其他潜在独特性的干扰。
在图5a中,实验者展示了选定位置在嵌入空间中的时间模式。虽然时间基元的基本形状在实现之间得以保留,但仔细观察会发现形状存在可变性。例如,C1的周期数量和循环频率可能有所不同,C4在强斜率之前可能显示出不同的平台。这些结果进一步证明了在输入空间上运行的CNN特征提取器具有非线性不变性。
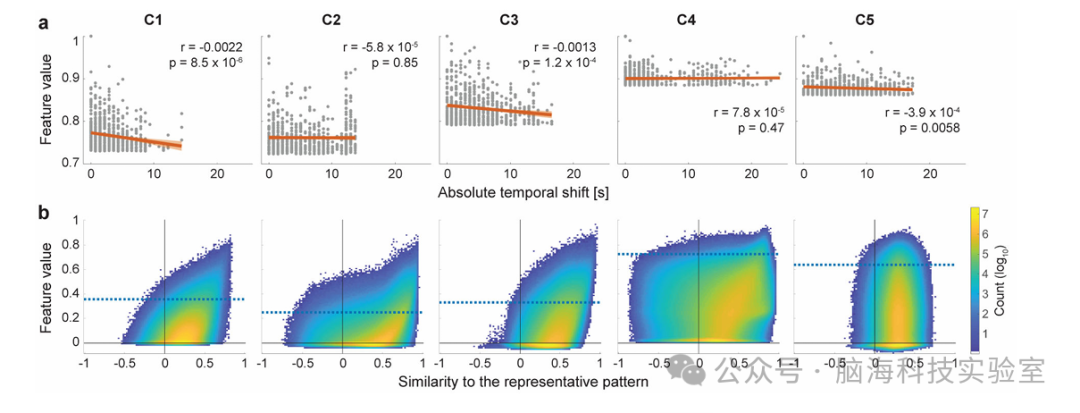
图4.特征提取器的不变性针对时间基原始的时间调制
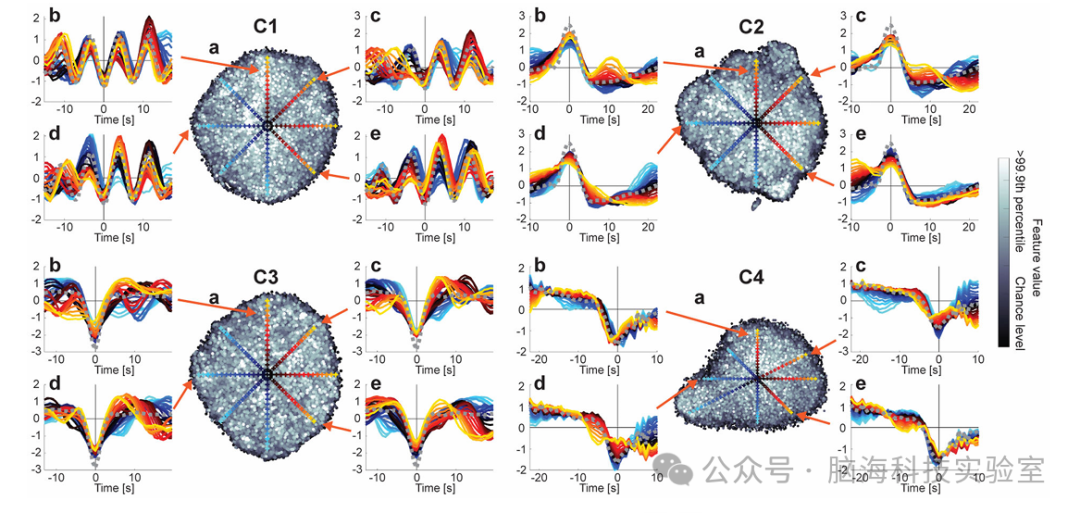
图5时间基元调制的可视化
图5展示了时间基元的非线性调制特性及其在输入空间中的变化。具体来说,通过将时间基元的实现(realizations)嵌入到二维空间中,并展示了这些实现的时间模式,揭示了CNN对时间基元的非线性调制的敏感性以及这些调制的多样性。
3.4时间基元的空间共现
为了揭示时间基元作为rfMRI信号中内在时间变化共同激活的基本元素,实验者利用二维嵌入技术可视化了它们的空间共现模式分布(见图6)。通过将时间基元的空间分布嵌入到二维空间中,揭示了这些时间基元在不同脑区的共现关系及其与已知静息态网络(RSNs)的关联。涵盖了默认模式网络(DMN)、后默认模式网络(PDMN)、侧默认模式网络(LDMN)、背注意网络(DAN)、额顶网络(FPN)、前扣带回网络(CON)、运动网络(MOT)和视觉网络(VIS)等。
具体而言,C2展现出广泛的空间共发生模式,与多个RSN高度相似,这表明C2可能是驱动这些静息态网络动态变化的主要因素。C3的共发生模式则较为局限,主要集中在FPN和VIS,偶尔涉及DMN,暗示其代表额顶控制网络和视觉网络的活动。C4的空间模式主要为任务阳性模式(包括DAN、MOT、FPN),并显著涉及DMN,结合其时间模式,C4似乎反映了任务相关激活(或努力失活)的持续存在,而非C2和C3所代表的瞬态活动。值得注意的是,这种持续的任务活动在静息状态下也自发出现。最后,C1虽与RSN相关的共发生模式多样,但空间焦点较少。总体上,各组件间某些共发生模式的重叠表明,与单个网络对应的时间模式并非总是相同,而是受到其他潜在大脑活动的调节,这再次印证了处理过程的非线性特性。
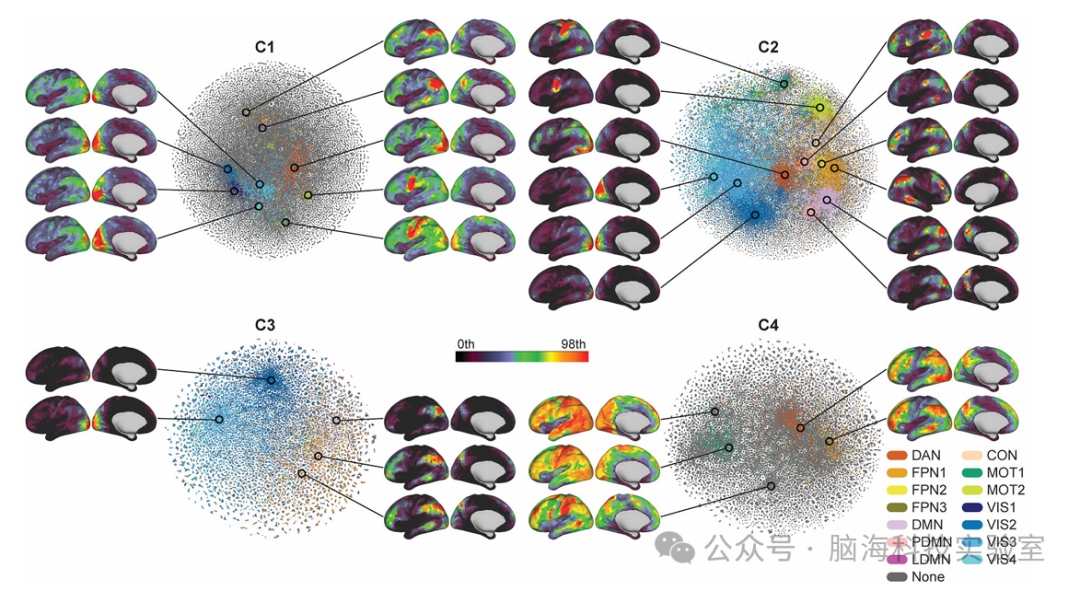
图 6时间基元的自发空间共现模式
散点图显示了静息状态期间时间基元的空间共现模式的二维t-SNE嵌入(有关t-SNE分析的详细信息)。颜色表示通过常规线性sICA获得的最相似的RSN;DAN(背侧注意力网络)、FPN(额顶叶网络)、DMN(默认模式网络)、PDMN(后 DMN)、LDMN(外侧 DMN)、CON(扣带回盖网络)、MOT(运动和体感网络)和VIS(视觉网络)。相似性通过Pearson相关测量,阈值为 0.35;灰点与小于0.35的RSN具有相似性。空间模式显示了嵌入空间上共现模式的一些示例,这些模式是通过在某些位置上取500个数据点的局部平均值获得的。仅显示左半球,但C2的FPN除外,因为它显示右主导模式。
3.5任务引起的时序基元
为了进一步研究时间原型,实验者评估了任务条件引起的时间模式和空间位置的实现(图7和8),通过将基于rfMRI训练的特征提取器应用于tfMRI数据。实验者发现,尽管有些任务具有独特的模式(特别是运动和语言任务,这可能是由于它们独特的区块设计),但在许多任务条件下,它们的时间模式和位置是一致的。C4在任务中始终出现在两个相对独特的时间点;任务开始后10.9 ± 2.1秒(15.1 ± 2.9体积)和任务块结束后6.4 ± 1.9秒(8.8 ± 2.6体积;语言任务除外,因为该任务块的长度变化较大,所以后者的峰值不明显)。这两个峰值的实现都显示出长时间的平台期,并略有下降趋势,这与众所周知的由区块设计范式引起的血氧水平依赖(BOLD)响应相似。其空间位置在后一个峰值与相应任务的效果大小图有明显的相似性,而在较早的峰值则分散在其他(与任务无关)区域。这些结果表明,C4代表了由持续任务在与任务相关的区域引起的BOLD响应,以及来自基线的与其他与任务无关的区域的扩展/努力去激活。
C3在许多任务中也表现出一致的峰值。首先,它出现在任务块结束附近(图7和8中的2b);时间和空间模式与效果大小图相似,表明它们代表了在与任务相关区域的块响应结束时出现的瞬态活动。C3还在许多任务条件开始后不久出现另一个峰值(图7和8中的2a),其时间模式与接收域响应(RRF)相似。其空间位置跨任务保持一致;主要位于FPN和视觉区域,这与静息状态时的共存模式一致(图6)(语言任务中的语言相关区域除外)。这些结果表明,在tfMRI期间,C3组织了FPN的负BOLD活动,以及在任务开始时刺激相关区域的血液动力学反应。C3和C4在视觉区域上的空间重叠可以通过HRF中刺激和任务相关成分之间的平衡调制来解释;即,在某些区块中,C3(刺激相关)占主导地位,而在其他区块中,C4(任务相关)占主导地位。
C2在许多任务中也表现出一致的峰值。首先,它出现在任务块结束附近(图7和8中的2b);时间和空间模式与效果大小图相似,表明它们代表了在与任务相关区域的块响应结束时出现的瞬态活动。C2还在许多任务条件开始后不久出现另一个峰值(图7和8中的2a),其时间模式与RRF相似。其空间位置跨任务保持一致;主要位于DMN相关区域、运动区域和视觉区域;这些区域中的许多都被发现与呼吸变化引起的信号重合。这些结果表明,C2既代表了与神经活动相关的反应,也代表了与刺激开始时的呼吸伪影同步的呼吸伪影。
尽管C1看起来不太具有时间特异性,但现在许多任务中的区块开始时明确地显示出周期性峰值。峰值的间隔约为7.2秒(10个体积),这与C1代表模式的周期一致。这意味着C1在任务区块期间出现在多个时间点,开始时与区块开始略微同步,然后逐渐失去同步。相应实现的空间位置在与任务无关的广泛区域上分散,这表明其生物基础与特定任务类型的相关性较小。
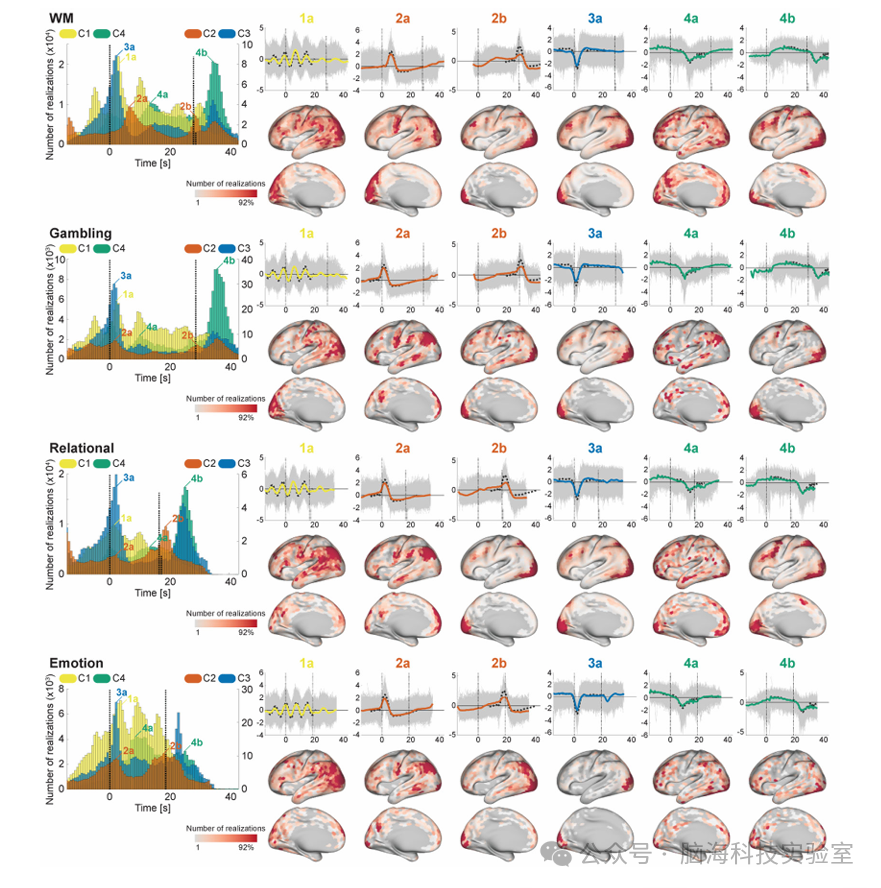
图7:任务条件下实现的时间和空间位置
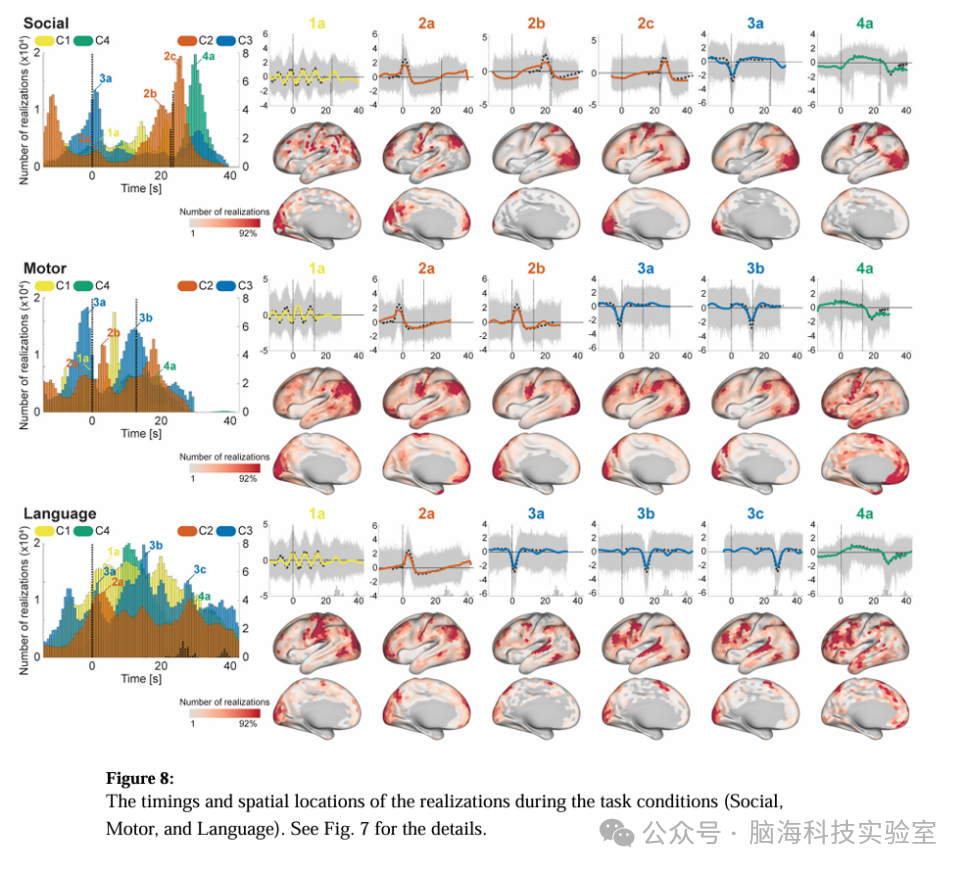
图 8在任务条件(Social、Motor和Language)期间实现的时间和空间位置
3.6时间基元与行为特征之间的关系
实验者通过分析每个时间基元在不同受试者中的平均活动水平,并将其与多种行为特征进行相关性分析,揭示了时间基元与认知能力、任务表现等行为特征之间的潜在联系。(图9a)。尽管这些相关性并不强,但一些特征与成分显示出显著的关系。C1与工作记忆相关的特征有显著相关性。C2和C3与流体智力测度相关;C3还与语言表现相关。
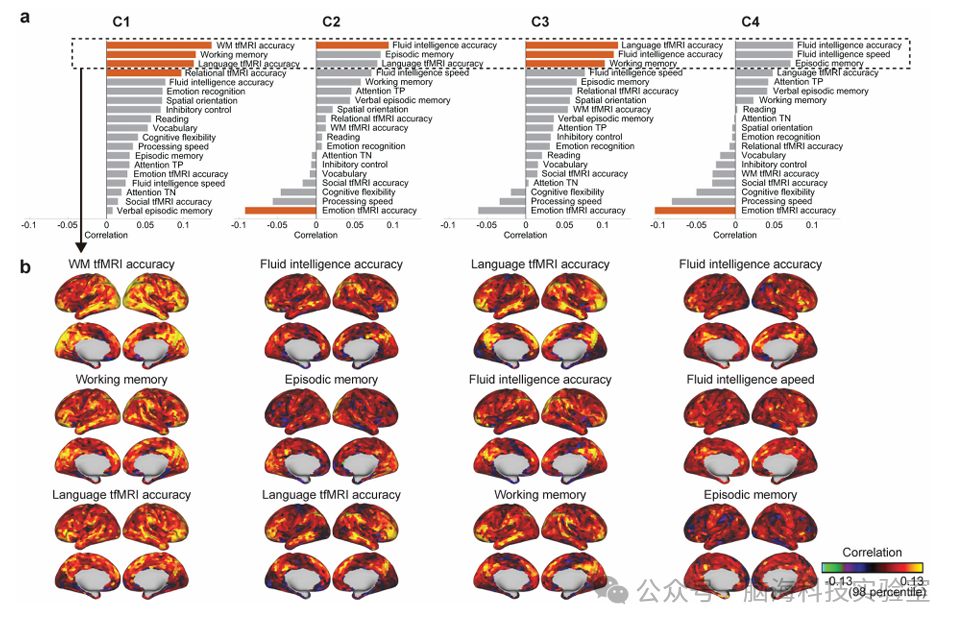
图9时间基元与受试者行为特征的关系
4讨论
(1)意义
时间基元是静息态与任务态动态网络的共同基础,支持“任务活动嵌入静息态”假说。
非线性特征提取对捕捉复杂调制至关重要,传统线性方法(如线性sICA)无法实现。
(2)局限性:
空间分割依赖:需预定义功能分区,可能影响结果稳定性;
成分数量选择:需权衡分解粒度与可解释性;
计算成本:CNN训练耗时(25小时),限制大规模应用。
总结
本文创新性地将深度学习与非线性ICA结合,提出LSCL框架,首次从fMRI中提取出具有行为意义的局部时间基元,为动态脑网络研究提供了新工具。其核心贡献在于:通过无监督学习解决非线性ICA的不可辨识问题,揭示时间基元作为跨状态(静息/任务)的神经动态单元。不足在于对预分割的依赖及计算复杂度,未来可通过自适应分割或轻量化网络改进。总体而言,LSCL为理解脑动态机制开辟了全新视角,是计算神经科学领域的重要进展。
END


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









