






 研究揭示了雷西诺受体选择性激动剂贝沙罗汀在p53缺失、MMTV-erbB2和C3(1)/SV40T抗原转基因小鼠乳腺癌模型中抑制肿瘤生长的作用。通过转录组分析,发现贝沙罗汀调节了涉及细胞周期、凋亡和代谢途径的基因表达,如蛋白质生物合成、三羧酸循环和氧化磷酸化的相关基因。这些发现表明,不同原癌基因途径来源的乳腺肿瘤可能对同一种化疗预防药物有不同的反应,并提出了氧化磷酸化诱导作为贝沙罗汀化疗预防效果的新机制。
研究揭示了雷西诺受体选择性激动剂贝沙罗汀在p53缺失、MMTV-erbB2和C3(1)/SV40T抗原转基因小鼠乳腺癌模型中抑制肿瘤生长的作用。通过转录组分析,发现贝沙罗汀调节了涉及细胞周期、凋亡和代谢途径的基因表达,如蛋白质生物合成、三羧酸循环和氧化磷酸化的相关基因。这些发现表明,不同原癌基因途径来源的乳腺肿瘤可能对同一种化疗预防药物有不同的反应,并提出了氧化磷酸化诱导作为贝沙罗汀化疗预防效果的新机制。
RXR-selective rexinoids inhibit the proliferation of normal, pre-malignant and malignant breast cells suppressing mammary tumor development in MMTV-erbB2, p53-Null, and C3(1)/SV40 T-antigen transgenic mice models [[6, 7] and Medina et al., unpublished]. The chemopreventive effects of bexarotene are likely due to transcriptional modulation of genes related to repression of cell proliferation and stimulation of apoptosis and cell differentiation [19].
In order to identify rexinoid-regulated biomarkers, we generated six mouse SAGE libraries corresponding to mammary gland samples from control and bexarotene treatment from three transgenic mouse mammary cancer models: p53-Null [10], MMTV-erbB2 [8] and C3(1)/SV40 T-antigen [9]. This resulted in the sequencing of 360,000 tags (60,000 tags per library), thus monitoring the behavior of more than 20,000 transcript tags. Our statistical analyses revealed 236 transcripts differentially regulated by bexarotene treatment in mammary epithelium from p53-null background, 283 transcripts in mammary gland from the MMTV-erbB2 model, and 290 transcripts in the C3(1)/SV40 T-antigen transgenic mice mammary model (Figure 2A; see Additional file 1). Table 1 shown the most highly bexarotene deregulated transcripts from each transgenic mice mammary cancer model (Fold change ≥ 7; p < 0.01).
Table 1 Most highly deregulated transcripts in mammary gland induced by bexarotene treatment on each transgenic mice mammary cancer model (Fold change ≥ 7; p < 0.01).
Figure 2
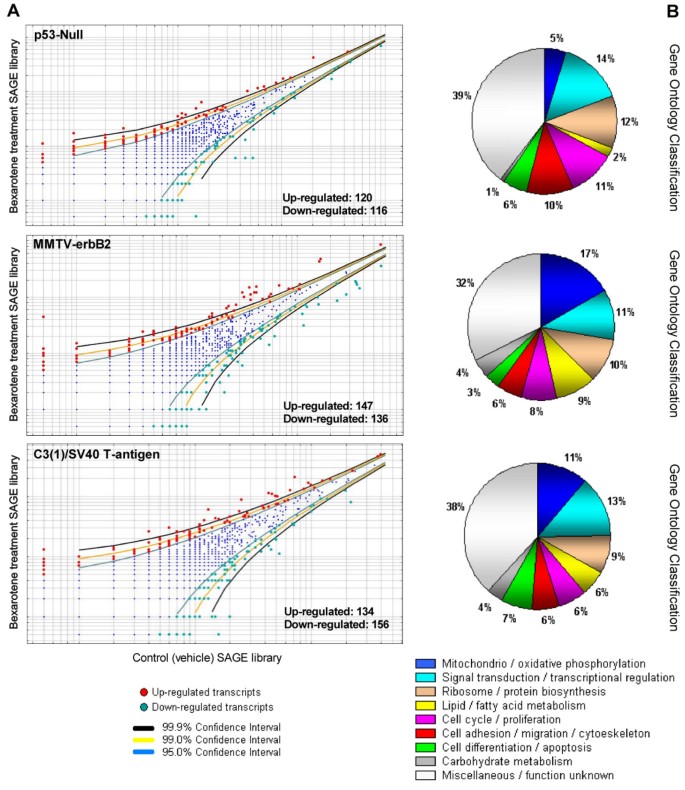
Deregulated transcripts in mammary gland by systemic treatment with bexarotene in the three transgenic mice mammary cancer models. A. Scatter-plot representation of differentially expressed genes between bexarotene treated mice and vehicle control SAGE libraries (p < 0.05). B. Gene ontology (GO) classification of bexarotene induced differentially expressed transcripts on mammary gland from the different transgenic models. Relative representation of the deregulated transcripts with specific GO term annotations related to biological processes or molecular function.
In order to identify co-occurring differentially expressed genes among the three transgenic mice analyses, we performed an inter-model comparison between the above-described SAGE datasets (Figure 1). Among the three mice mammary models, a total of 711 transcripts were identified as deregulated by the rexinoid bexarotene treatment. Eighty-nine genes were identified in more than one mammary cancer model (Figure 3A; see Additional file 2). Interestingly, nine of these 89 genes were deregulated by bexarotene in mammary gland tissue from all three transgenic models: Muc15 (Mucin 15), Cdo1 (Cystein dioxygenase 1), Rps8 (Ribosomal protein S28), Rps27 (Ribosomal protein S27), Rps24 (Ribosomal protein S24), Hspa5 (Heat shock 70 kD protein 5), Csrp1 (Cysteine and glycine-rich protein 1), Npm1 (Nucleophosmin 1), and Cycs (Cytochrome c somatic). Gene Ontology annotation of the 89 deregulated genes that were common in any two models showed that approximately 18% of the transcripts are involved in tricarboxylic acid cycle/oxidative phosphorylation, 14% are related to signal transduction/transcriptional regulation, 14% are related to protein metabolism and 12% are related to cell proliferation/differentiation and apoptosis.
Figure 3
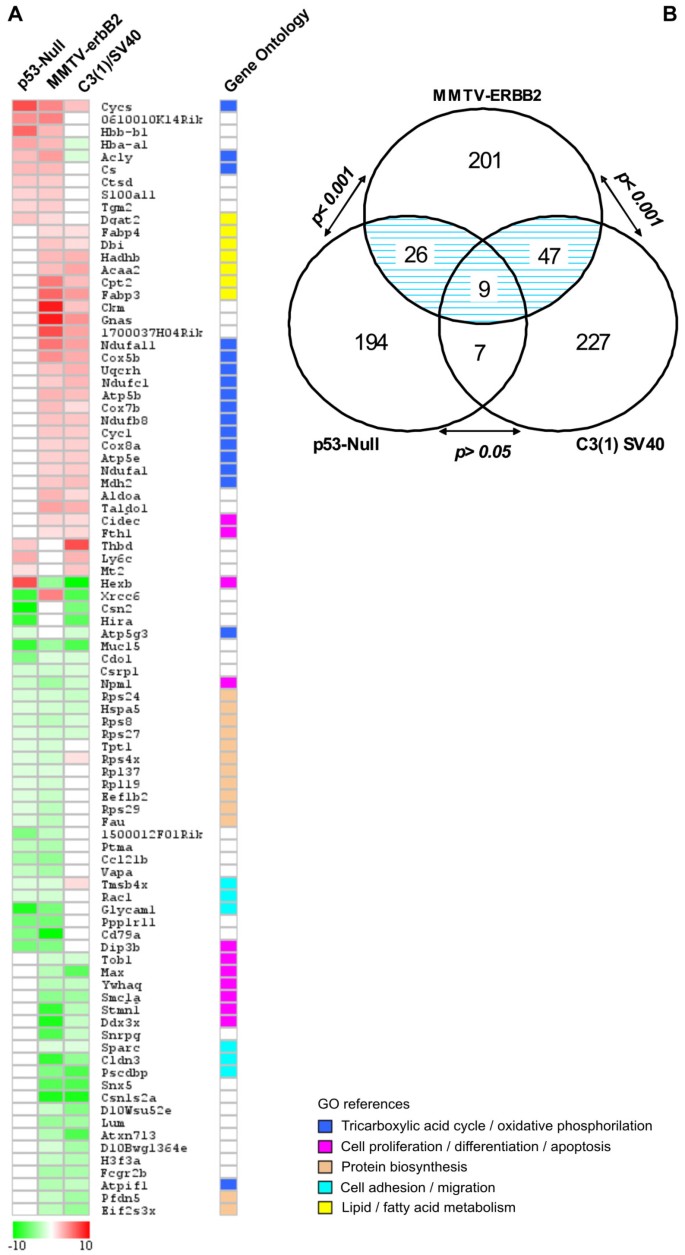
Co-occurring differentially expressed genes among transgenic mouse mammary models. Eighty-nine genes were identified as modulated in more than one transgenic mice model. A. Heat map of the 89 deregulated transcripts. Color scale at the bottom depicts the approximate fold change in expression for each transcript and library relative to control mammary gland. Negative fold change (transcripts with decreased expression in bexarotene treated animals) is represented in green, and positive fold change (transcripts with overexpression in bexarotene treated mice) is represented in red. B. Statistical comparison between MMTV-erbB2 vs. p53-null and MMTV-erbB2 vs. C3(1)/SV40 T-antigen transgenic mice models showing a highly significant number of overlapping genes (p < 0.001). The number of overlapping genes between p53-null and C3(1) SV40 models it is not statistical significant (p > 0.05).
A probabilistic analysis showed that 56 genes were co-deregulated in MMTV-erbB2 and C3(1)/SV40 T-antigen mice models, representing a non-random significant number of overlapping genes based on normal approximation to the binomial distribution (p < 0.001) (Figure 3B). Thirty-five genes were identified as co-deregulated in MMTV-erbB2 and p53-null mice models (p < 0.001). The set of 16 genes overlapping between p53-null and C3(1)/SV40 T-antigen were not statistical significant, i.e. the overlapping could be simply by chance (p > 0.05) (Figure 3B). In other words, it appears that a better correlation was observed between MMTV-ErbB2 with the other two models, than between p53-null and C3(1)-SV40 tag transgenic mouse mammary gland models. These data suggest that mammary tumors derived from different primary oncogenic pathways could respond differently to the same chemoprevention agent. In addition, these results indicates that transcripts modulated by bexarotene in the MMTV-ErbB2 mammary gland share almost all the common features among the transgenic mouse models analyzed. As mentioned above, we have previously shown that bexarotene suppresses mammary tumor development in the MMTV-ErbB2, p53-null and C3(1)-SV40 tag transgenic mouse mammary gland models [6, 7]. Interestingly, the specific response of these three transgenic mouse mammary models to bexarotene treatment varies with the genetic background assessed. For instance, the bexarotene treatment is much more effective against MMTV-ErbB2 induced mammary tumors than against C3(1)-SV40 or p53-null mammary tumors [Medina et al., unpublished]. In the MMTV-ErbB2 mammary gland, bexarotene reduced tumor incidence by 75% and lengthened median tumor latency from 234 days to over 420 days [7]. However, in the p53-null and C3(1)-SV40 mammary gland where p53 or p53/Rb activities are affected respectively, bexarotene treatment showed modest chemoprevention activity. Both these molecules exert primary functions downstream of the CDKs, loci of targets activity. In this sense, human breast cancer is a complex disease caused by dysregulation of many different oncogenes, tumor suppressor genes and growth factor pathways. The MMTV-ErbB2, p53-null and C3(1)-SV40 tag mouse mammary gland cancer models are valuable tools for the elucidation of the mechanisms of mammary tumorigenesis [3]. However, it is important to recognize that no one model represents the heterogeneity of human breast cancer.
We present in Figure 4 a protein-protein interaction network associating the common core of non-random bexarotene modulated genes across transgenic mouse mammary models. The graph was generated employing the STRING on-line resource based on high confidence data related with 'co-expression/co-ocurrence', 'experimental/biochemical data' and 'association in curated database/text mining' [17]. STRING is a comprehensive tool integrating protein association information with the capability to transfer known interactions from model organisms to other species (e.g.: from mouse to human orthology genes/proteins) based on predicted orthology of the respective proteins. The generated graph (Figure 4) indicates strong interactions among a set of 33 proteins transcriptionally modulated by bexarotene. Furthermore, the network architecture suggests the existence of two functional modules in this figure, involving the down-modulation of genes related with protein biosynthesis pathway, and up-modulation of genes related with tricarboxylic acid cycle/oxidative phosphorilation pathways.
Figure 4
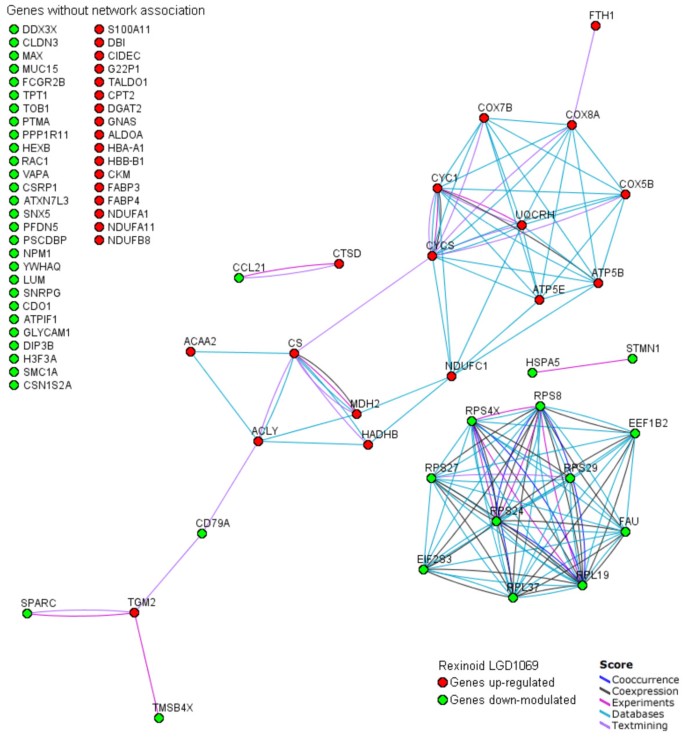
Graph of interactions among the common core of genes modulated by rexinoid bexarotene in the different mammary mice genetic models generated using database STRING. Genes without known interactions with other genes are listed in the left of the figure. In the network: links between proteins means the various interactions data supporting the network, colored by evidence type.
Protein biosynthesis signature
A common observation in cancer gene expression profiling is the systematic up-regulation of ribosomal genes among the most abundant transcripts in human and mouse mammary carcinomas compared with normal tissues [20–24]. The up-regulation of ribosomal genes was significantly correlated with variation in the cell doubling time in vitro, supporting the notion that these genes are up-regulated in relation to the increase of cell proliferation rate or growth rate during malignant transformation. Interestingly and in an opposite manner, bexarotene treatment cause in 'normal' mammary gland the down-regulated expression of more than 10 genes related to protein biosynthesis including numerous ribosomal proteins (Rpl19, Rpl37, Rps4x, Rps8, Rps24, Rps27, Rps29), Eef1b2 (Eukaryotic translation elongation factor 1 beta 2), Eif2s3x (Eukaryotic translation initiation factor 2), Fau (Finkel-Biskis-Reilly murine sarcoma virus) and Tpt1 (tumor protein, translationally-controlled 1). The inhibition of mRNA synthesis for genes encoding ribosomal proteins has been suggested as a mechanism that could reprogram the cancer cell to recover some of its normal functions in a tumor reversion process [25].
Tpt1 (also known as Tctp) encodes a GDP dissociation inhibitor protein of the translation elongation factor eEF1A [26]. The human TPT1 gene is overexpressed in cancerous cell lines compared with cell lines derived from normal tissues. Tuynder et al. (2002) demonstrated that the expression levels of TPT1 were strongly down-regulated at the mRNA and protein levels during tumor reversion/suppression. MCF7 and T47D cell lines transfected with Tpt1 siRNA showed a more organized ductal-like structures similar to those generated by down-regulation of β1 integrin [25]. Here we observed that bexarotene significantly dowregulated expression of Tpt1 in mammary epithelium.
Bioenergetics signatures
More than 50 years ago, Warburg proposed that malignant phenotype might be caused by a decrease in mitochondrial energy metabolism paralleled by increased glycolytic flux [27]. Increasing evidence is in line with this hypothesis suggesting a close link between metabolic and genetic changes observed during malignant growth [28, 29]. Recently it has been demonstrated that impaired bioenergetic function of mitochondria is a hallmark of carcinogenesis in breast, gastric, lung and oesophageal cancer [30, 31]. Moreover, Schulz et al. (2006) showed that induction of mitochondrial oxidative metabolism efficiently suppresses malignant growth in vitro and in vivo. Interestingly, we identified a systematic up-regulation of transcripts related to oxidative phosphorylation induced by bexarotene treatment in mammary gland (Figure 4). The transcripts commonly up-regulated by bexarotene treatment in at least two of the models were Atp5b (ATP synthase F1 complex beta subunit), Atp5e (ATP synthase F1 complex epsilon subunit), Cyc1 (Cytochrome c-1), Cycs (Cytochrome c somatic), Cox5b (Cytochrome c oxidase, subunit Vb), Cox7b (Cytochrome c oxidase subunit VIIb), Cox8a (Cytochrome c oxidase, subunit VIIIa), Ndufa1 (NADH dehydrogenase 1 alpha subcomplex), Ndufc1 (NADH dehydrogenase 1), Ndufb8 (NADH dehydrogenase 1 beta subcomplex 8), Ndufa11 (NADH dehydrogenase 1 alpha subcomplex 11) and Uqcrh (Ubiquinol-cytochrome c reductase hinge protein) (see Additional files 1 and 2). Consistent with a significant increase of oxidative phosphorylation enzymes, we observed that Atpif1 gene (ATPase inhibitory factor 1) was significantly down-regulated by bexarotene treatment in the MMTV-erbB2 and C3(1)/SV40 T-antigen transgenic mice models. In this sense, Isidoro et al. (2005) showed that down-regulation of ATPase β-F1 per se allowed the identification of a subgroup of breast cancer patients with significant worse prognosis. Finally, is important to note that mitochondrial oxidative phosphorylation is required for efficient execution of apoptosis. Cells which are unable to carry on oxidative phosphorylation have a resistant apoptotic phenotype [32]. Overall, these findings suggest the oxidative phosphorylation induction (prevention impaired bioenergetic function) as a novel mechanism of bexarotene's chemopreventive effects.
Fatty acid metabolic signature
Lipid metabolism and the intracellular transport of bioactive species is a critical component in the process by which these molecules continuously stimulate proliferation through interactions with nuclear receptors. Bexarotene treatment of MMTV-ErbB2 and C3(1)/SV40 transgenic mammary gland up-regulated various genes related with lipid/fatty acid metabolism (Figure 4) such as: Fabp3 (Fatty acid binding protein 3), Fabp4 (Fatty acid binding protein 4), Dgat2 (Diacylglycerol O-acyltransferase 2), Dbi (Diazepam binding inhibitor), Hadhb (Hydroxyacyl-Coenzyme A dehydrogenase), Acca2 (Acetyl-Coenzyme A acyltransferase 2), and Cpt2 (Carnitine palmitoytransferase 2). Interestingly, a family of cytoplasmic proteins known as FABPs mediates transport and utilization of lipids, and different FABP types have been implicated in control of cell proliferation and cancer progression. Recently, FABP1 and FABP2 were shown to be up-regulated in breast cancer cell lines while FABP3 and FABP4 were down-regulated in breast cancer cells [33]. Moreover, FABP4 is a marker protein for differentiated mammary gland that is expressed only in normal lactating cells and not in tumor mammary cells. Transfection of cDNA clone of FABP4 into MCF7 cells results in growth inhibition and lower tumorgenicity in nude mice [34].
Cell proliferation and apoptosis signatures
Rexinoid bexarotene down-regulated several genes related with cell cycle/proliferation in mammary gland from the different transgenic mice models (Figure 2A; see Additional files 1 and 2).
Among this functional group we find Npm1 (also known as Nucleophosmin/B23) protein that belongs to a nuclear chaperone family of phosphoproteins that take part in various cellular processes such as cell proliferation and transformation [35]. Human NPM1 is overexpressed in various tumors types, and it has been proposed as a marker for gastric, colon, ovarian and prostate carcinomas [35]. NPM1 overexpression promotes cell survival in several cell types through the inhibition of distinct pro-apoptotic pathways [36]. We detected a systematic down-regulation of Npm1 gene in mammary gland from Bexarotene treated mice on the three models studied (average fold change = -2.6; p < 0.01). Interestingly, proteomic analyses identified NPM1 as a protein associated with acquired estrogen-independence in human breast cancer cells [37]. Moreover, down-regulation of NPM1 mRNA delay cell-cycle progression and the entry into mitosis [38], whereas NPM1 overexpression decreases the sensitivity of human leukaemia cells to retinoic-acid-induced differentiation and apoptosis [39, 40].
Another gene in this category includes Stmn1 which encodes an 18 kDa cytosolic phosphoprotein (also known as Stathmin 1 or Oncoprotein 18) that is regulated during cell cycle by transcriptional and posttranscriptional mechanisms. STMN1 overexpression has been demonstrated at mRNA and protein levels in a significant proportion of human breast carcinomas (about 30%) [41]. Moreover, STMN1 overexpression was correlated with the loss of ERα and with histological grade III breast carcinomas. STMN1 has been suggested as a key regulator of the cell division through its influence on microtubule dynamics. We identified a statistical significant decrease of Stmn1 expression (average fold change = -5.4; p < 0.05) caused by bexarotene treatment in mammary gland from MMTV-erbB2 and C3(1)/SV40 T-antigen mice. Interestingly, we previously demonstrated that mouse Stmn1 and human homologue STMN1 genes are overexpressed in invasive breast carcinomas by northern and real time RT-PCR analyses [24].
Numerous studies have shown that down-regulation of p27Kip1, an inhibitor of cyclin-dependent kinase, is associated with poor prognosis in many cancers such as: breast, colorectal, prostate, and lung carcinomas. We previously detected the overexpression of CDC28 protein kinase regulatory subunit 1B (Cks1b) in human and mouse mammary tumors [24]. Interestingly, rexinoid bexarotene strongly down-regulated Cks1b expression in the MMTV-erbB2 model (Fold change = -10.0; p = 0.006) (see Additional file 1). Human CKS1B functions as an important adaptor of SCF Skp2 ubiquitin ligase and facilitates SCF Skp2 targeting of the cell proliferation inhibitor p27 (Kip1) for ubiquitination and subsequent degradation [42]. It was also suggested that CKS1B may be involved in p21 degradation in a similar fashion [43]. Overexpression of CKS1B has been observed associated to poorly differentiated tumors (histological grade III) and with the loss of ER/PR status [Slotky et al., 2005]. In addition, CKS1B overexpression was strongly and independently associated with poor overall survival in human breast cancer [44].
On the other hand, bexarotene treatment up-modulated two apoptotis related genes (Cidec and Cycs) in 'normal' mouse mammary gland from two of the models (Cidec) and in all three models (Cycs) (Figure 3). Cidec (also known as Fsp27) encode a novel family member of the cell-death-inducing DFF45-like-effectors (CIDEs) [45]. Although, its well known that DFF45 is a subunit of the DNA fragmentation factor that is cleaved by caspase-3 during apoptosis [46]. The molecular mechanism by which Cidec induces apoptosis remains to be elucidated.
Cell adhesion and invasion signatures
During their metastatic conversion, epithelial cells acquire the ability to invade the surrounding tissues and later disseminate to secondary organs mostly via lymphatic vessels. Epithelial cell adhesions, including intercellular (junctional) and cell-extracellular matrix adhesions, are critical to the maintenance of structural integrity, polarity, and cell-cell communication. We detected a significant decrease in Cldn3 (Claudin 3) (Average fold change = -6), Glycam1 (Glycosylation dependent cell adhesion molecule 1) (Average fold change = -7), Pscdbp (Pleckstrin homogy Sec7 binding protein) (Average fold change = -6) gene expression modulated by bexarotene treatment among transgenic mice models. The Claudin genes (Cldn) encode a family of proteins important in epithelial cell tight junction, which are critical to the maintenance of cell polarity and permeability [47, 48]. Most Claudin genes appear with decreased expression in cancer however CLDN3 and CLDN4 genes have been found frequently up-regulated in ovarian, breast, prostate and pancreatic cancers [49–52]. Recently, has been suggested that Claudins may be envolved in survival and invasion of cancer cells [48]. We detected down-regulation of Cldn3 gene in mammary gland from bexarotene treated mice in the MMTV-erbB2 and C3(1)/SV40 models. The role of Gycam1 and Pscdbp genes in breast cancer progression remains unknowns.

















 1万+
1万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









